Alternatív tulajdonságok a genetikában. Genetikai szakkifejezések szójegyzéke
Szótár genetikai kifejezések
Autolízis - a szövetek, sejtek vagy részeik önemésztése saját enzimek hatására állatokban, növényekben és mikroorganizmusokban.
Az autoszintézis ugyanazon szülői forma kromoszómáinak konjugációja egy távoli hibridben.
Adaptív (indukált) enzimek - enzimek, amelyek szintézisének sebessége a létezés körülményeitől függően változik. A szintézis szabályozása genetikai szinten történik induktorok hatására, amelyek lehetnek a megfelelő szubsztrátok és metabolitok, valamint hormonok (az E. coli las régiójának enzimei, β-galaktozidáz, permeáz, acetiláz; galaktóz szolgálhat). szubsztrátként, és az izopropil-β szolgálhat induktorként -D-tiogalaktozid, IPTG).
Adapter - transzfer RNS (tRNS) molekula, amely a transzláció során aminosavakat szállít a hírvivő RNS-hez (mRNS).
Az additív gének olyan polimer gének, amelyek azonos hatással vannak a fenotípusra, de összegző hatást fejtenek ki.
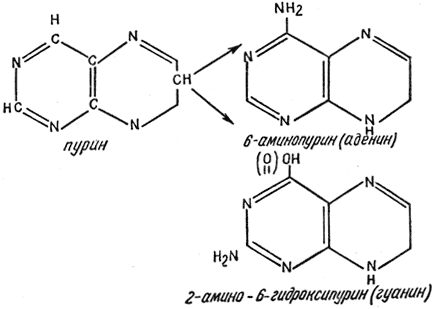
A nitrogéntartalmú bázisok a nukleinsavakat alkotó bázisok. Két fő típusa van: pirimidin (uracil, timin, citozin) és purin (adenin, guanin).
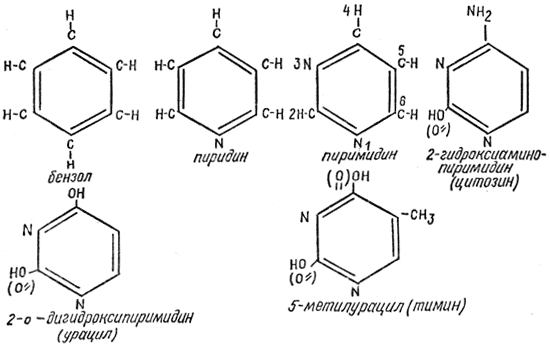
A pirimidin nukleotidok pirimidingyűrűkön alapulnak:
Aminosav aktiválás - aminoacil-tRNS képződése. Ez a folyamat magában foglalja az aminosav tényleges aktiválását és az aktivált aminoacil-maradék átvitelét a tRNS-molekulára, ami aminoacil-tRNS-t képez. Mindkét reakciót az egyes aminosavra specifikus aminoacil-tRNS szintetáz (vagy aktivációs enzim) katalizálja. Az aminosavat az ATP aktiválja úgy, hogy egy AMP-maradékot kapcsol a karboxilcsoportjához. A keletkező aminosav-adenilát az aktivációs enzimhez kötődik: AA + ATP + tRNS → aminoacil-tRNS + AMP + oldat.
Az akridinek (akridin színezékek) - akridinnarancs, akriflavin, proflavin, valamint az alkilezett akridinek (ICR-170, ICR-191) erős mutagén hatást fejtenek ki, eltolódást okozva a leolvasási keretben.
Az enzim aktív helye egy specifikus hely a felszínen, amelynek köszönhetően az enzim szubsztrát-specifitást mutat. Az egy polipeptidláncból álló enzimeknek egy aktív helyük van. A β-galaktozidáz molekulának négy aktív centruma van – egy-egy a négy azonos polipeptidlánchoz, amelyek a szerkezetének kialakításában vesznek részt. Az aktív centrum jelenléte az enzim háromdimenziós konformációjának következménye, mivel az azt alkotó aminosavmaradékok a polipeptidlánc különböző részein helyezkednek el.
Alkilezőszerek - mutagének (etil-metánszulfonát, nitrozometil-karbamid stb.), amelyek képesek a nitrogéntartalmú bázisok megváltoztatására szénhidrogén-maradékok - etil és metil - bejuttatásával.
Az allél a gén egyik lehetséges állapota. A gén szerkezetében a mutációk vagy a heterozigóták belső rekombinációi következtében két mutáns allél esetében bekövetkező bármilyen változás a gén új alléljainak megjelenéséhez vezet. Az alternatív tulajdonságokat meghatározó génpárt allelomorf párnak, magát a párosítási jelenséget pedig allelomorfizmusnak vagy allelizmusnak nevezzük. Különböztesd meg a homoalléleket és a heteroallélokat. Homoallélek (izoallelák) - allélek, amelyek közötti különbségek csak egy helyre vonatkoznak. A heteroallélok olyan allélek, amelyek különböző helyeken különböznek egymástól, és képesek rekombinációkat létrehozni az intragénikus átkelés miatt.
Több allél - sok gén esetében nem kettő, hanem több vagy akár sok allélállapot is ismert. Többszörös allélizmus esetén egy adott génnek mindig csak egy allélja van jelen egy ivarsejtben vagy spórában, és ennek a génnek két (azonos vagy eltérő) allélja mindig jelen van a diploid szervezetek sejtjeiben. A több allél hasítása mindig monohibrid marad.
A mutagén allélspecifitása – a mutagén azon képessége, hogy allélelváltozásokat hordozó mutánsok reverzióját idézze elő.
Allopoliploid. Lásd Mutáció, Amphidiploid.
Allosyndes - különböző kromoszómák konjugációja szülő nyomtatványok távoli hibridben.
Az alloszterikus hatás egy olyan jelenség, amelyben egy enzim vagy szabályozó fehérje konfigurációja és biológiai aktivitása megváltozik azáltal, hogy egy kis molekulatömegű anyagot - egy effektort (represszor fehérje és laktóz) kapcsolnak hozzá.
Az allofén szervezetek olyan kiméra szervezetek, amelyek genetikailag különböző embriók blasztomereinek kombinációjából fejlődnek ki.
Az alloenzimek (allotípusok) ugyanazon lókusz (gén) különböző alléljai által kódolt enzimek. A Drosophilában az alkohol-dehidrogenáznak két allotípusa ismert, amelyek szubsztrátspecifitásukban különböznek egymástól, az A-forma és a B-forma. Az A-forma jobb, mint a B-forma az etanol, izopropanol és ciklohexanol feldolgozásakor. Magasabb hőmérsékleten a B-forma stabilabb és aktívabb, mint az A-forma, és van néhány szelektív előnye vele szemben. Az allo-enzimek jelenléte növeli a populációk polimorfizmusát, ami növeli életképességét. Lásd az izoenzimeket.
A borostyánszupresszorok olyan mutáns gének, amelyek tRNS-t kódolnak, és képesek felismerni (szignifikáns) borostyánkodonokat (UAG).
Az ambivalencia (ambivalens fágok) egy olyan jelenség, amelyben a T4 fág egyes rII-mutánsai nem tudnak növekedni egyes K-baktériumtörzseken, másokon viszont igen. Az ambivalens mutánsok nonszensz mutációt tartalmaznak az rIIA génben, ami miatt a tőle jobbra található rIIB régió transzlációja nem történik meg. A K permisszív törzsei nonszensz szuppresszorokat (nonszensz mutáció-szuppresszorokat) tartalmaznak, aminek eredményeként a nonszensz kodon értelmesként kerül beolvasásra, és a transzláció normálisan megy végbe.
A riboszóma aminoacil centruma - a riboszóma 50S helye, amelyhez az aminosavat hordozó tRNS (aminoacil-tRNS) kapcsolódik, ha ennek a tRNS-nek az antikodonja megfelel az mRNS kodonjának, amely a Ebben a pillanatban aminoacil központjában található.
Az amniocentézis kis mennyiségű magzatvíz szúrással történő eltávolítása, amelyben az embrionális sejtek lebegnek. A sejteket mesterséges tápközegen tenyésztik, és citogenetikai és biokémiailag vizsgálják. A mutációk és az embrió nemének korai felismerésére használják.
Génamplifikáció - a genom egyes részeinek szelektív sokszorosítása. Az rRNS génreplikáció egy speciális típusa, amikor egy részük elhagyja a kromoszómát a nukleáris nedvhez, a magmembrán közelében helyezkedik el, és ott folytatja autonóm replikációját. Átírás után nagyszámú Az rRNS-molekulák bejutnak a citoplazmába, és riboszómák képzésében használják fel őket. Kétéltűek, rovarok, egyes puhatestűek petesejtekben, növényi portokok bélésrétegében figyelhető meg.
Amfidiploid - olyan organizmus, amely interspecifikus hibridizáció alapján jön létre, és két diploid kromoszómakészlettel rendelkezik. Sok interspecifikus növényhibrid azért steril, mert a különböző szülőktől kapott kromoszómák nem konjugálnak a meiózis során, véletlenszerűen oszlanak el, ezért a meiózis termékei nem életképesek. De ha egy ilyen hibridben a kromoszómák száma megduplázódik, a meiózis normálisan megy végbe, mivel egy ilyen tetraploid hibridnek két kromoszómakészlete van mindkét szülőfajból; a meiózis során a kromoszómák homológjaikkal konjugálnak, és helyesen térnek el a keletkező ivarsejtekbe. Az amfidiploidok megszerzése egy módszer a távoli hibridek termékenységének helyreállítására.
Az amphimixis a növények és állatok ivaros szaporodási módja, amelynek során az apai és az anyai ivarsejtek összeolvadásával új szervezet jön létre.
Legközelebbi szomszéd frekvencia analízis – az újonnan szintetizált komplementer DNS-szál bázisszekvenciája azonosságának bizonyítására szolgál.
A nukleinsavban négyféle bázis található. Ezért a dinukleotidok 16 kombinációja lehetséges. A. Kornberg egy módszert javasolt annak meghatározására, hogy a 16 kombináció mindegyike milyen gyakorisággal fordul elő a vizsgált DNS-ben. Ehhez négy dezoxiribonukleozid-5′-trifoszfátot veszünk. Az egyik például, amely a pentóz 5' szénatomjához kapcsolódik, 32 P-vel van jelölve az α pozícióban:
A nyíl a polimerizáció utáni törés helyét jelzi.
A polimerizációt követően a képződött DNS hidrolízisen megy keresztül az 5'-C dezoxiribóz és a foszfát közötti foszfodiészter kötéseket specifikusan felbontó enzimek hatására, ami dezoxiribonukleozid-3'-monofoszfátok képződéséhez vezet:
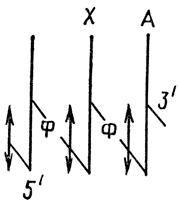
Az eredetileg az αATP tulajdonában lévő 32P jelölés αCMP formájában, azaz a nukleozid 3'-monofoszfátban található, amely a polimerben az adenil-nukleotid mellett volt.
A kapott négy dezoxiribonukleozid-3'-monofoszfátban meghatározzuk radioaktivitásukat, és kiszámítjuk, hogy a polimerben milyen gyakorisággal foglal helyet az adenin mellett, azaz négy dinukleotid szekvencia, TfA, CfA, GfA, AfA gyakoriságát számítjuk ki. . Hasonló kísérletet végzünk egy másik jelölt nukleotiddal, például dGTP-vel (32P 2), és hidrolízis után négy másik nukleotid gyakoriságát határozzuk meg. TfG, CfG, AFG, GfG szekvenciák, majd a többi trifoszfáttal - dCTP, dTTP (32 P). Az elemzés lehetővé teszi számunkra, hogy a következő következtetéseket vonjuk le.
1. Mind a 16 dinukleotid szekvencia megtalálható a DNS-ben, és gyakoriságuk a DNS jellemzője.
2. A reakcióban szereplő DNS-ben és az újonnan szintetizált DNS-ben a legközelebbi szomszédok gyakorisága megegyezik, azaz az első DNS valóban templátként szolgál a szintézisben:
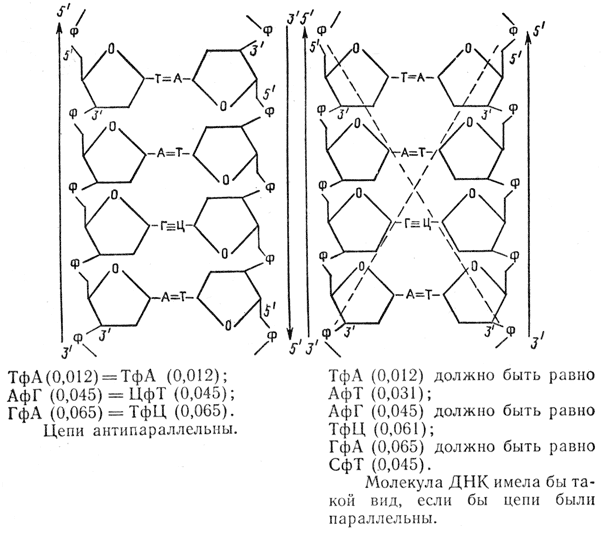
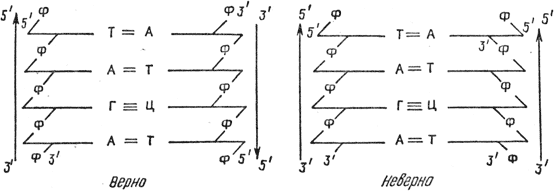
3. Két újonnan képződött DNS-szál antiparallel, mivel az elméleti dinukleotid-frekvenciák egybeesnek a kísérleti adatokkal:
Egy ismeretlen genotípusú fenotípusú domináns organizmus keresztezésének elemzése recesszív szervezet. A hasítás meglététől vagy hiányától függően a domináns organizmus következő genotípusait különböztetjük meg - heterozigóta és homozigóta. A két fenotípusos osztályra (1(A) + 1(a) való felosztás bizonyítékul szolgál egy, a tulajdonságot szabályozó gén jelenlétére. A négy fenotípusos osztályra bontás a keresztezett formák dihibriditásának bizonyítéka.
A bázisanalógok a purinok és pirimidinek, amelyek eltérnek a nukleinsavak szokásos nitrogénbázisaitól. Beépülhetnek a nukleinsavakba, és mutációk indukálásához vezethetnek.
Az androgenezis az organizmusok olyan szaporodási formája, amelyben a hím sejtmag, amelyet a hímivarsejtek juttatnak be a petesejtbe, részt vesz az embrió fejlődésében, a női mag pedig nem vesz részt.
Antigén - antitest reakció - egy antigén specifikus kötődése a megfelelő antitesthez, ami immunkomplex kialakulásához vezet.
Az antigének olyan anyagok, amelyeket a szervezet idegenként érzékel, és specifikus immunhatást váltanak ki, amely kölcsönhatásba léphet e válasz termékeivel - antitestekkel (immunglobulinokkal) és immunocitákkal in vivo és in vitro egyaránt. Az antigén tulajdonságok minden élő szervezet makromolekuláris összetevőivel rendelkeznek.
Az antikodon a tRNS-molekula három nukleotidból álló szakasza, amely felismeri a megfelelő három nukleotidból álló szakaszát (kodon), amely a megfelelő aminosavat kódolja a hírvivő (messenger) RNS-ben. A transzláció során ezt a kodont a kodon és az antikodon komplementaritása miatt csak egy specifikus tRNS ismeri fel, így biztosított az aminosavak beépülésének szigorú sorrendje és sorrendje a növekvő polipeptidláncba.
Az antimutagének olyan tényezők, amelyek csökkentik a mutációk gyakoriságát. Ide tartoznak a különféle kémiai természetű vegyületek - ciszteamin, kinakrin, néhány szulfonamid, propionsav és galluszsav származékai.
Az anti-termináció a triptofán és a triptofán operon csillapítója kölcsönhatásának leírására használt kifejezés. A triptofán jelenléte a tápközegben az r-mRNS vezető szekvenciája transzkripciójának befejezéséhez vezet, a triptofán hiánya a teljes triptofán-operon transzkripcióját, azaz az anti-terminációt határozza meg. Lásd Csillapító.
Az apogámia a növényekben előforduló apomixis egyik formája. Lásd: Apomixis.
Az apomixis egy szervezet szaporodása, amelyet nem kísér szexuális folyamat. Szűkebb értelemben másodlagos ivartalan szaporodás, amelyben a korábbi szaporodási szakaszok megsértése miatt az embrió megtermékenyítés nélkül fejlődik. Attól függően, hogy az ivar (tojás) vagy a vegetatív sejt hoz létre új szervezetet, az apomixis két fő formáját különböztetjük meg - a partenogenezist és az apogámiát.
Aposporia – kiesés életciklus a sporulációs folyamat növényei, és ezért a haploid forma.
Csillapító - a trp operonban az O lókusz és a trp E gén kezdete közötti régió, amely részt vesz a triptofán szintézis szabályozásában.
Az arginin egy aminosav, amely a fehérjék, különösen a protaminok (akár 85%) és a hisztonok része.
Auxotróf mutáció - olyan mutáció, amely a szervezet életéhez szükséges egyik vagy másik összetett szerves anyag szintézisének képességének elvesztéséhez vezet. Például egy bakteriális mutáns csak komplett táptalajon kolonizál, minimális táptalajon nem nő.
Outbreeding - keresztezés vagy keresztezési rendszer ugyanazon faj nem rokon formáival. Ebben az esetben egyazon fajtához vagy fajtához (fajtákon belüli vagy fajtán belüli keresztezés) és különböző szervezetekhez (fajták közötti vagy fajták közötti keresztezések) tartozó szervezetek is keresztezhetők. Ha nem rokon egyedeket keresztezünk, a káros recesszív mutációk heterozigótákká válnak, és az első generációs hibridek gyakran életképesebbnek és betegségekkel szemben ellenállóbbnak bizonyulnak, mint szüleik, és megnövekedett termékenységük. A kitenyésztés segítségével különféle értékes tulajdonságok kombinálásával új állat- vagy növényfajta jön létre. Mivel a kitenyésztés eredményeként olyan hibridek jönnek létre, amelyek a kombinációs variabilitás következtében jobb és rosszabb tulajdonságkombinációkkal is rendelkezhetnek, ezért a kívánt formák kiválasztásának mindig követnie kell a keresztezést.
Autoimmun kivételek - autonóm izogén plazmidok replikációjának elnyomása azonos nevű plazmid jelenlétében, amely már a replikáció szigorú ellenőrzése alá került. A mérsékelt égövi fágokban ez immunitás az elfojtás általi felülfertőződéssel szemben, az F-faktorban pedig a felületes kizárás.
A DNS autokatalitikus funkciója a DNS azon képessége, hogy szabályozza saját replikációját a benne lévő genetikai információ miatt, amely biztosítja a komplementer molekula szintéziséhez szükséges összes folyamat és mechanizmus végrehajtását:

Autoszómák – a kétlaki állatok, növények és gombák sejtjeiben található összes kromoszóma, a nemi kromoszómák kivételével. A betűvel vannak jelölve. A nőstény Drosophila kromoszómaképlete: 6A + XX; férfi - 6A + XY.
A bakteriofágok olyan vírusok, amelyek baktériumokban szaporodnak.
A bakteriocinek bizonyos baktériumok által termelt specifikus fehérjék, amelyek gátolják az azonos fajhoz vagy rokon baktériumfajhoz tartozó más törzsek sejtjeinek létfontosságú aktivitását. Az E. coli által termelt bakteriocinokat colicinnek nevezik. A legtöbbet tanulmányozott Col E1, Col E2 és Col E3 plazmidok az enterobaktériumok természetes populációinak 20%-ában találhatók meg. Az összes colicint a baktériumsejtek felszínén található speciális receptorok ismerik fel: egy részük behatol a sejtbe, mások a sejtmembrán módosulásával hatnak. Tehát a Col E1 gátolja a celluláris foszforilációt, a Col E2 egy DNS endonukleáz, a Col E3 megszakítja a fehérje bioszintézist, változást okozva a riboszómális 30S alegységben (a 16S-pRNS részleges lebomlása figyelhető meg).
Keresztezi. Lásd visszakeresztezés.
iker módszer- az egyik módja annak, hogy az ikrek összehasonlító elemzésével tisztázzuk az öröklődés és a környezet relatív szerepét a tulajdonságok változékonyságában. Az ikrek genetikai vizsgálatához az alábbi kritériumok alapján kell meghatározni a típusukat: 1) az egypetéjű ikreknek azonos neműeknek kell lenniük; 2) az egypetéjű ikreket konkordanciával (hasonlósággal), a testvérikreket - sokféleképpen, beleértve a vércsoportokat is, diszskordanciával (hasonlósággal) kell jellemezni; 3) egypetéjű ikreknél a reciprok szövettranszplantáció, mint az autotranszplantáció, nem végződhet kilökődéssel. Ikreknél ez az immunológiai inkompatibilitás miatt lehetetlen. Az egypetéjű ikerpárnak azonos genotípusa van, ami lehetővé teszi a környezetnek a tulajdonságok kialakulásában betöltött szerepének megismerését (például ikernevelés különböző körülmények között). Az azonos környezetben élő két ikertípus összehasonlítása feltárja az öröklődés szerepét a tulajdonság kialakulásában. Ugyanakkor szem előtt kell tartani, hogy az ember számára a környezet nemcsak fizikai tényezők, hanem társadalmi feltételek is. Az iker módszer lehetővé teszi számos betegség, például skizofrénia, tuberkulózis és angolkórra való örökletes hajlam meghatározását.
Vándorgének. Lásd transzpozonok.
A vektor egy autonóm módon replikálódó genetikai struktúra, amely segítségével a benne lévő gént át lehet vinni a megfelelő genomváltozásba. A vektor körkörös bakteriális plazmidok (R-plazmidok, λ fág genom a genom egy részének deléciójával, F-faktor). A gének emlőssejtekbe történő átviteléhez az onkogén SV40 vírust használják, amely néha pszeudovírusokká alakul, amelyben a vírusfehérje kapszidjában (héjában) nem vírus DNS található, hanem annak a sejtnek egy DNS-fragmentuma, amelyben a vírus replikálódott. .
Génkölcsönhatás - sok gén hatása egy tulajdonságra. Mivel minden fehérje aminosavakból áll, a fehérjeszintézisben nemcsak az elsődleges szerkezetét szabályozó gén vesz részt, hanem maguk az aminosavak szintézisét biztosító gének is.
A Drosophila-ban több mint 50 gén mutációját írták le, amelyek befolyásolják a szem színét. A szervezet összetett tulajdonságai, mint például az életképesség és a termékenység, szintén nagyszámú géntől függenek. A gének kölcsönhatása az úgynevezett génegyensúly példáján mutatható be, vagyis a nemi kromoszómák és az autoszómák arányának a kétlaki élőlények nemi jellemzőire gyakorolt hatása. A fejlődést a női oldalra irányító gének (a Drosophilában) főként az X kromoszómán, a hím oldalra fejlődést irányító gének pedig az autoszómák különböző helyein (második és harmadik kromoszóma) helyezkednek el. A gének kölcsönhatása a genetikai információ megvalósításának minden szakaszában megnyilvánul. Hagyományosan a génkölcsönhatások következő formái különböztethetők meg: 1) egy gén elsődleges termékének (polipeptidjének) hatása egy másik gén transzkripciójára; 2) különböző gének transzkripciós termékei közötti reakciók; 3) azon ontogenetikai folyamatok különböző láncainak keresztezése, amelyek meghatározásában ezek a gének részt vettek.
Fenotípusos szinten a következő típusú génkölcsönhatások különböztethetők meg: 1) komplementáció, 2) neoplazma, 3) episztázis, 4) kriptoméria, 5) polimerizmus.
A kontextus befolyása a rekombinációra a gén egy bizonyos régiójában a rekombináció gyakoriságának a nukleotid-összetételtől való függése, ami a linearitás megsértéséhez vezet a kéthelyes keresztezéseken alapuló rekombinációs térképek felépítésében.
Nem tervezett szintézis - DNS-szintézis, amelyet annak károsodása indukál.
A mutagének intragén specifitása a mutagének azon képessége, hogy direkt mutációkat indukálnak, amelyek a megnyilvánulás lokalizációjában vagy természetében különböznek (világos vagy homályos manifesztációval), valamint interallélikus komplementációra vagy szuppresszálhatóságra való képességükben. A mutagén affinitását tükrözi a gén egy adott régiójához. A mutagének specifitását a "forró pontok" megnyilvánulása érzékeli.
belső szupresszorok. Lásd: Váltáscsillapítók olvasása.
Visszakeresztezés - valamelyik szülői forma első generációjának hibridjével vagy genotípusában hasonló formával keresztezzük.
Forgásszimmetria – a nukleotidszekvenciák ugyanúgy olvashatók, ha 180°-kal elfordítják őket (inverzió). Ezek a helyek lac O-val vannak jelölve, ahol represszor kötőhelyként szolgálnak, és BAC kapcsolódási helyet képeznek a laktóz operon promoterben.
Haploidia - saját fajának kromoszómakészletének jelenléte a sejtben.
A helikázok (helikázok) letekeredő fehérjék (destabilizálják a DNS-hélix hélixet), és megakadályozzák az egyszálú fragmensek újraegyesülését. Akár 200 helikáz fordul elő a replikációs villában, és mindegyik molekula 8-10 nukleotidból álló komplexet alkot. A kötés nem specifikus a bázisok tekintetében. A helikázokat fágokból és baktériumokból izoláltuk (fd fd - fehérje 5, fág T4 - génfehérje 32).
A hemizigóta állapot egy olyan állapot, amelyben az egyednek csak egy adagja van bizonyos génekből, ezért nem lehet sem homozigóta, sem heterozigóta. Az X kromoszómán elhelyezkedő gének hemizigótái a hím kétszárnyú rovarok, emlősök és nőstény madarak.
A gén az öröklődő információ szerkezeti egysége; a genetikai anyag funkcionálisan oszthatatlan egysége. A gén egy DNS-molekula szakasza (egyes vírusok esetében RNS), amely egy polipeptid, egy transzport vagy riboszomális RNS-molekula elsődleges szerkezetét kódolja, vagy kölcsönhatásba lép egy szabályozó fehérjével.
A sejtben szintetizálódnak polipeptideket kódoló szerkezeti gének, amelyek meghatározzák az rRNS, tRNS szerkezetét, és olyan akceptor gének, amelyek a replikációban, transzkripcióban és a génaktivitás szabályozásában részt vevő egyes enzimek specifikus kapcsolódási helyeiként szolgálnak.
Genealógiai elemzés - bizonyos tulajdonságok öröklődési mintáinak elemzése a törzskönyv (genealógia) összehasonlítása alapján. Az elemzést abban az esetben végzik el, ha ismertek a közvetlen törzskönyvek - az örökletes tulajdonság (proband) tulajdonosának ősei anyai és apai ágon több generációban, vagy a proband leszármazottai több generációban. genealógiai módszer orvosi genetikai konzultációkon használják.
A genetikai analízis egy szervezet örökletes tulajdonságainak tanulmányozására szolgáló módszerek összessége. A genetikai elemzés fő módszerei közé tartozik a tenyésztési, hibridológiai, citogenetikai, populációs, molekuláris genetikai, mutációs és ikerelemzés.
A genetikai vagy populációs homeosztázis a panmiktikus populáció azon képessége, hogy a változó környezeti feltételek mellett fenntartsa a genotípusos szerkezet viszonylagos stabilitását és integritását. A homeosztázis mechanizmusai közé tartozik a populáció egyensúlyi állapotának fenntartása a genetikai gyakoriságok tekintetében a Hardy-Weinberg képlet szerint, a heterozigótaság és a polimorfizmus fenntartása, a mutációs folyamat bizonyos sebességének és irányának fenntartása.
Genetikai terhelés - a populáció örökletes változékonyságának része, amely meghatározza a kevésbé alkalmazkodó egyedek megjelenését, akik szelektív halálnak vannak kitéve a természetes szelekció folyamatában. A populáció alkalmasságát csökkentő genetikai terhelés két összetevőből áll: egy spontán mutációs folyamatból, amely leggyakrabban káros következményekkel jár, valamint a gének hasadása, kombinálása következtében új, kevésbé adaptált genotípusok megjelenéséből. Mivel azonban a mutagenezis és a gének rekombinációja a genotípus hasznos változásainak forrása, ami a variabilitás mobilizációs tartalékát jelenti, a genetikai terhelés „fizetésként” működik a faj további fejlesztésének lehetőségéért.
A genetikai kód egyetlen rendszer, amely az élő szervezetekre jellemző nukleinsavmolekulák örökletes információit nukleotidszekvencia formájában rögzíti. A genetikai kód fő tulajdonságai: 1) triplet - minden aminosavat három nukleotid kódol (UUU - fenilalanin, CCC - prolin, CAU - hisztidin); 2) megszakítás nélküli - az egy tripletthez (kodonhoz) tartozó nukleotidok nem szerepelnek a szomszédos hármasokban és 3) degeneráció - egy aminosavat több hármas (prolin - CCC, CCA, CCU, CCG) kódolhat, amelyeket ekvivalensnek neveznek. A kétértékű kodonok egy csoportját kódsorozatnak nevezzük. A metioninnak és a triptofánnak egy-egy kódoló hármasa van - AUG, UGG. A többi aminosav több hármasnak felel meg, például prolin, hisztidin - négy, arginin, leucin, szerin - hat hármas. Az UAA, UAG, UGA hármasok terminációs kodonokként (nonszensz kodonok) szolgálnak, amelyek a transzláció végét jelzik.
Tegyen különbséget szisztematikus és nem szisztematikus degeneráció között. Az ilyen degenerációt szisztematikusnak nevezzük, ha az ekvivalens kodonpárokban a 3'-terminális pozícióban purinok (A és G) vagy pirimidinek (U és C) különböznek egymástól, a többi eset a nem szisztematikus degeneráció példája.
Genetikai anyag - sejtkomponensek, amelyek szerkezeti és funkcionális tulajdonsága biztosítja az örökletes információk tárolását, megvalósítását és továbbítását a vegetatív és ivaros szaporodás során. A genetikai anyag univerzális tulajdonságokkal rendelkezik minden élőlény számára: diszkrétség, folytonosság, linearitás és viszonylagos stabilitás. A diszkrétség egy gén, kromoszómák és genom létezése. A diszkrétség e gének allélkészletének formájában nyilvánul meg, amely megfelel a kapcsolódási csoportnak - kromoszómának; a genomnak megfelelő kapcsolódási csoportok halmaza. Folytonosság - a kromoszóma fizikai integritása, számos gén egymáshoz való kapcsolódásában nyilvánul meg a kromoszóma-átrendeződések eredményeként feltárt különféle helyzethatásokban, valamint az operon poláris és szabályozó mutációiban. Linearitás - a genetikai információ rögzítésének egydimenzióssága, amely egy kapcsolódási csoporton belül egy bizonyos génszekvenciában vagy egy génen belüli helyeken mutatható ki. Relatív stabilitás - a változatok megjelenése és megőrzése a genetikai anyag reprodukciója során, mutációs variabilitás formájában nyilvánul meg. A konvariáns reduplikáció képessége a genetikai anyag szaporodása és módosítása, amelyet a megváltozott változatok szaporodása követ.
A mutagén génspecifitása - a mutagén jellemzője, amely megmutatja, hogy képes változásokat okozni bármely adott génben.
Gén egyensúly. Lásd: Génkölcsönhatás.
Genom - egy adott típusú szervezet haploid kromoszómakészletére jellemző összes gén összessége; fő haploid kromoszómakészlet.
Genotípus - egy organizmus genetikai (örökletes) felépítése, egy adott sejt vagy organizmus összes örökletes hajlamának összessége, beleértve a gének alléljait, a kromoszómák fizikai kapcsolatának természetét és a kromoszóma-átrendeződések jelenlétét. Vannak homozigóták, amelyek mindkét homológ kromoszómában azonos allélokkal rendelkeznek homológ lókuszokon (AA, aa), és ugyanazokat az ivarsejteket alkotják egy adott gén különböző alléljeivel.
Genotrófok - olyan organizmusok, amelyek a táplálkozás változása következtében megváltoztak. A genotrófok képződése a kromoszómák különböző részeinek amplifikációján alapulhat.
A mutátor gének olyan gének, amelyek a genom mutabilitási szintjét szabályozzák.
A szupresszor gének olyan gének, amelyek képesek elnyomni más gének fenotípusos expresszióját.
Heteroallélok. Lásd allél.
Heterogametikus ivar - olyan nem, amely kétféle ivarsejtet képez, amelyek nemi kromoszómákban különböznek egymástól. Az XX-XY és XX-X0 rendszerben a hím heterogametikus, a ZZ-ZW és ZZ-Z0 rendszerben a nőstény heterogametikus. Az XX-XY típusú ivarmeghatározás a legtöbb organizmusban megtalálható - emlősökben, kétszárnyú rovarokban, halakban; XX-X0 - néhány poloska számára. A ZZ-ZW rendszer és származéka ZZ-Z0 kevésbé elterjedt. Nőstény heterogametiát találtak szamócában, lepkékben, egyes halakban, kétéltűekben és szinte minden madárban, a növények között - az eperben.
Heterogén. Lásd merozigóta.
heteroduplex modell. Lásd: Heterozigóta belső.
Belső heterozigóta - a nukleotidok közötti kovalens kötések kialakulása miatt korlátozott heterozigóta régióval rendelkező DNS-molekulák jelenléte a vegetatív alapban T-egyenetlen fágok:
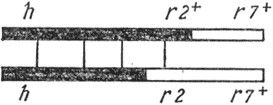
Ez a heterozigóta homozigóta hr2 + r7 + és rekombináns hr2r7 + DNS molekulákra hasad a félig konzervatív replikáció során.
Komplex heterozigóta - bizonyos növényfajokban, például az Oenothera lamarckianában, két különböző genom jelenléte. Az egyes haploid kromoszómakészleteken belüli kromoszómák átrendeződéséből adódó speciális citológiai mechanizmusnak köszönhetően az összes kromoszóma összekapcsolódik egymással, így a meiózis során nem egyesülnek újra, és a két kromoszómakészlet közül az egyik bejut az egyes ivarsejtekbe - vagy gaudenek, vagy velánok. . Az ilyen heterozigótaság állandóságát a kiegyensúlyozott letálisok tartják fenn, amelyeket heterozigóta állapotban más gének normál alléljai elnyomnak.
A populáció heterozigótasága a populációk mutációkkal való telítettsége, amely örökletes variabilitásuk tartalékát képezi, amely lehetővé teszi, hogy genetikai szerkezetének megváltoztatásával alkalmazkodjon a feltételekhez. A populáció egyedeinek heterozigóta állapota biztosítja annak adaptív plaszticitását. Ezen túlmenően a heterozigóták életképessége nagyobb, mint a homozigótáké, szélesebb a reakciósebessége, vagyis nagyobb az adaptív képességeik, mint a homozigótáké, ami szelektív előnyt biztosít számukra.
Heterózis - megnövekedett vitalitás, termékenység az első generációs hibridekben, nem rokon keresztezéssel.
Heteroimmun fágok - a 434 és λ fágok, egymáshoz képest heteroimmunok, mivel mindegyikük a második fágra lizogén baktériumokon nő, azaz mindegyik érzéketlen egy másik fág represszorának hatására.
A DNS heterokatalitikus funkciója a DNS azon képessége, hogy a benne található genetikai információknak köszönhetően szabályozza az összes létfontosságú szerves molekula - aminosavak, polipeptidek, vitaminok stb. - szintézisét:

A kromoszóma heteromorfizmus olyan jelenség, amikor a homológ kromoszómák morfológiailag különböznek egymástól (például mutációk következtében).
Heterokromatin. Lásd: Kromatin.
Heterotall klónok - szexuális folyamat élesztőben, amely két haploid sejt fúziójából áll, amelyek két szexuális típushoz tartoznak. Vannak heterotallikus és homotallikus klónok. Az első esetben a sejtek bármelyik szexuális típushoz tartoznak, a második esetben a sejtek mindkét szexuális típus keverékei. A sütőélesztőben az egyik vagy másik szexuális típusú (a és a) sejtek képződésének szabályozása a következőképpen történik: az egyik kromoszómában két „csendes” gén, az 1-es és a 2-es gén található. Közöttük a „szexuális lókusz” " PTL, amelyben vagy a gén egy kópiája van jelen 1-es (α nemi típus), vagy a 2. gén kópiája (a nemi típus), azaz a nemi szabályozás ebben az esetben strukturális gének transzponálásával történik. A homotalizmust meghatározó HO gén részt vesz egy ilyen transzpozíció megvalósításában. Amikor osztódik, az élesztőtörzs heterotallikussá válik, azaz egyik vagy másik nemi típus rögzül.
A heterotrófok olyan szervezetek, amelyek exogén szerves anyagokat használnak szénforrásként.
Heterofil - különbségek a levelek alakjában, méretében és szerkezetében ugyanazon a növényen.
Hibrid - genotípusosan eltérő organizmusok (sejtek) genetikai anyagának kombinálása, azaz hibridizáció eredményeként kapott organizmus.
A szomatikus sejtek hibridizálása olyan technikák rendszere, amely lehetővé teszi különféle mesterséges környezetben tenyésztett szervezetek (egerek és patkányok, egerek és emberek, emberek és csirkék, csirkék és élesztők) sejtjeinek fúzióját. Az ilyen hibrid sejtekben olyan gének kölcsönhatása figyelhető meg, amelyek más módon nem kombinálhatók. Mivel az osztódás során az egyes kromoszómák elveszhetnek, azonosítható a sejt fenotípusára vagy az enzimek szintézisére gyakorolt hatásuk. A szomatikus sejtek hibridizációjának segítségével számos gén lokalizációját határozták meg a humán kromoszómákban.
A hibridológiai elemzés a tulajdonságok öröklődésének jellegének elemzése keresztezési rendszer segítségével. Hibridek megszerzéséből és azok továbbfejlesztéséből áll összehasonlító elemzés több generáción keresztül.
Nukleinsavak hidrolízise. Lásd: Nukleinsav-szekvenálás.
Gynandromorfok - egyedek, amelyek testének egy része női, egy része férfi szerkezetű. A Drosophilában a gynandromorfok általában azért képződnek, mert a megtermékenyített XX petesejt osztódásának különböző szakaszaiban az egyik blasztomer mindkét X kromoszómát (a női testrész), a másik pedig egy X kromoszómát (a hím) kap. rész). A test minden részén megjelenhetnek jelek az X kromoszóma génkészletének megfelelően. A gynandromorfizmust két haploid mag jelenléte okozhatja egy megtermékenyítetlen petesejtben, és a polispermia (rovaroknál gyakori) miatt a megtermékenyített magok eltérő ivari kromoszómákkal rendelkezhetnek.
A kétértelmű levelezés hipotézise (az angol wooble hipotézisből), vagy a swing hipotézis. E hipotézis szerint az mRNS és a tRNS közötti kölcsönhatás során a riboszómán csak a kodon első két bázisa fog szükségszerűen standard komplementer párokat alkotni az antikodon megfelelő nukleotidjaival. Kölcsönhatás során a kodon harmadik nukleotidja párokat alkothat az antikodon különböző bázisaival. A harmadik pozíció kétértelmű megfeleltetése lehetővé teszi egy pár G-U, A-I (inozinsav, amely purin bázist tartalmaz az antikodonban - hipoxantin) kialakulását. A tRNS-antikodon harmadik nukleotidja az mRNS-kodonokban lévő bázisok bizonyos tartományát képes felismerni. Példa a kodon és az antikodon közötti kétértelmű megfeleltetésre az élesztő alanin tRNS-e, amelynek a CGI antikodonja három alaninkodonra - GCC-re, GCC-re és GCA-ra - reagál. A negyedik GCH alanin kodon felismeréséhez azonban van egy második alanin tRNS, amely a CGU vagy CHC antikodonnal rendelkezik.
Szekvencia hipotézis. Ezen elmélet szerint a génelemek szekvenciája határozza meg a polipeptidláncban lévő aminosavak sorrendjét.
A girázok (girázok) a topoizomeráz II, amelyek negatív szuperspirálozást okoznak a körkörös DNS-molekulákban egyetlen kötés megszakításával, elforgatásával és egy kötés helyettesítésével. Az E. coli-ban például a girázt a gyr A és gyr B gének szabályozzák.
A hisztonok a növényi és állati sejtek magjában található fehérjék. Gazdag arginin- és lizinmaradékban, amelyek meghatározzák lúgos tulajdonságaikat. Molekulatömege 11200-21000. A sejtmagokban DNS-komplexként van jelen, és fontos szerepet játszik a csomagolásában. A kromatinban a hisztonok a száraz tömeg 25-40%-át teszik ki. A hisztonok stabilizálják a kromatin szerveződését, a nukleinsavak szintézisének szabályozásában az egyik láncszemként szolgálnak, jelentősen növelik a sejtmembránok permeabilitását a makromolekuláris vegyületek számára. A hisztonok fajspecifikussága gyengén kifejeződik. A disszociált nukleoszómák rekonstrukciója elvégezhető különböző élőlényekből vett hisztonok részvételével. Lásd: Nukleoszóma.
A hollandi tulajdonságok olyan tulajdonságok, amelyek csak a férfi vonalon keresztül öröklődnek az őket szabályozó gének Y kromoszómában való elhelyezkedése miatt, például a lábujjak közötti bőrhártya jelenléte, az embernél szőrös fülek.
A homeotikus mutációk olyan mutációk, amelyekben egy másik szerv helyett másik fejlődik ki, különösen a Drosophilában, végtagok képződhetnek az antenna rudimentumából.
A homeosztázis a biológiai rendszerek azon képessége, hogy ellenálljanak a változásoknak, és fenntartsák az összetétel és a tulajdonságok dinamikus relatív állandóságát.
Homoallelák. Lásd allélek.
Homogametikus ivar – olyan ivar, amely a nemi kromoszómákhoz képest azonos ivarsejteket képez. Lásd a heterogametikus szexet.
A homológ kromoszómák olyan kromoszómák, amelyek ugyanahhoz a párhoz tartoznak. A diplomid szervezeteknek annyi kromoszómapárja van, ahány különböző kromoszóma van a megfelelő haploid halmazban.
Homológia - a szervek megfeleltetése az organizmusokban különböző típusok filogenetikai kapcsolatuk miatt.
homotallikus klónok. Lásd a heterotallikus klónokat.
A mutációk "forró pontjai" megnövekedett mutációs rátával rendelkező helyek (a T4 fágszegmensben A - 300 mutáció, B - 500 mutáció). A „forró pontok” eloszlásának jellege az alkalmazott mutagéntől függően változhat.
A kapcsolódási csoport olyan gének összessége, amely ugyanazon a kromoszómán található, és ezért összekapcsolt módon öröklődik. A különböző kromoszómákon elhelyezkedő, azaz különböző kapcsoltsági csoportokhoz tartozó gének egymástól függetlenül öröklődnek. A kapcsolódási csoportok száma megegyezik a kromoszómák haploid számával.
Mutációs nyomás - az allélgyakoriság arányának változása egy populációban a gén közvetlen és fordított mutációinak eltérő valószínűsége miatt. Ennek eredményeként megnő annak az allélnek a gyakorisága, amelynek irányában nagyobb gyakorisággal fordulnak elő mutációk. Ha a direkt és a reverz mutációk egyformán valószínűek, a mutációs nyomás megszűnik, és egyensúlyi állapot lép fel a populációban.
Osztások - egyfajta kromoszóma-átrendeződés, amelynek eredményeként a kromoszómagén belső szakasza kiesik. Lásd mutáció.
A hibás fágok olyan HFT-lizátumokból származó fágok, amelyek képesek transzdukcióra (például λ gal fág), amelyekben genomjuk 30%-át a bakteriális gal régió helyettesíti. Ennek eredményeként a gal defektív fágokból hiányzik néhány létfontosságú gén a λ genom h régiójában. Ezért nem hibás vagy aktívan lizogén Gal + transzduktánsok csak akkor nyerhetők, ha a nem lizogén Gal - baktériumokat a HFT lizátumból származó fággal fertőzték meg. A HFT-lizátum a λ-profág ultraibolya indukciójával jön létre lizogén Gal +/Gal-heterozigóták tenyészetében, ahol nagy a fertőzések száma, így a transzdukáló λ-gal-fággal fertőzött sejt egyidejűleg egy normális, nem fertőzött sejttel is megfertőződik. transzdukáló részecske.
A timin dimerizáció a polinukleotid láncon elhelyezkedő két szomszédos timinmaradék megkötése az egyik ultraibolya kvantum abszorpciója következtében. A dimerizáció a DNS kettős hélix másodlagos szerkezetének lokális felbomlásához és annak a génnek a funkciójának elnyomásához vezet, amelyben előfordult.
A kromatincsökkenés a genom egy részének kizárása az eukariótákban az embriogenezis vagy a protozoon makronukleuszának érése során.
Diploidia - a fajra jellemző két kromoszómakészlet jelenléte a sejtben.
Az eltérés az ikrek jeleinek különbözősége. Lásd iker módszer.
diszkrétség. Lásd a genetikai anyagot.
A disztális gének olyan gének, amelyek egymástól távol helyezkednek el a kromoszómán.
A hosszú távú módosítások olyan módosulások, amelyek több generációt érintenek a vegetatív vagy partenogenetikus szaporodás során.
A nemi differenciálódás az ontogenetikus szexuális jellemzők kialakulásának folyamata. Az állati embriók kezdetleges szexuálisan közömbös ivarmirigyei kettős természetűek. Ide tartozik a külső réteg - a kéreg, amelyből a női csírasejtek fejlődnek ki a differenciálódás folyamatában, és a belső réteg - a velő, amelyből a hím ivarsejtek fejlődnek. A differenciálódás során az ivarmirigy egyik rétege kifejlődik és a másik elnyomja. Az állatok nemi differenciálódási folyamatát a nemi szerv megfelelő rétegei, majd az ivarmirigyek által kiválasztott csírák határozzák meg. A magasabb rendű növények nemi differenciálódását a növényi hormonok - auxinok - befolyásolják.
A gének eltérő aktivitása - egy többsejtű szervezet minden sejtje azonos génkészleteket tartalmaz, de más idő a különböző szövetekben különböző gének működnek, aminek köszönhetően differenciálódás történik. A génaktivitás szabályozása különböző szinteken történhet: replikáció, transzkripció, transzláció. A replikáció szintjén történő szabályozás a jelenleg szükségtelen gének kópiák számának növelésével vagy csökkentésével történik. Lásd: Génamplifikáció.
A DNS-polimerázok nagy molekulatömegű polimerek, amelyek katalizálják a nukleotidok hozzáadását a DNS-lánc 3'-OH-végéhez.
A kiegészítő kromoszómák (B-kromoszómák) kisméretű, teljesen heterokromatikus kromoszómák, amelyek nem szerepelnek az adott fajra (rozs, kukorica, pocok, gyíkok, rovarok) jellemző standardkészletben. Nem minden egyedben találhatók meg, és a kromoszómakészlet véletlenszerű kiegészítései. A fenotípusokra gyakorolt hatás elhanyagolható. Sok B-kromoszóma hátrányosan befolyásolja az életképességet és a termékenységet.
Egy gén dózisa egy adott gén másolatainak száma a genomban.
A domének egy fehérje harmadlagos szerkezetének kialakulásának félig autonóm egységei. Röntgen-diffrakciós elemzéssel számos fehérjében megtalálható. Bemutatták a domének különféle kombinációinak (E. coli és N. crassa triptofán-szintetáz) örökletes rögzítésének lehetőségét. A tartományi együttműködés interallélikus komplementációt okozhat.
Dominancia (domináns allél, mutáció, tulajdonság) - az egyik szülő tulajdonságának túlsúlya az első generáció hibridjében. Az egyik allél másik feletti dominanciájának megnyilvánulásához a domináns allélnak kellő mennyiségű molekulaszintézist kell biztosítania, amelyek meghatározzák a génhatás specifitását, a recesszív allélnak pedig az inaktív molekulák szintézisét kell biztosítania. ezt a funkciót, és nem lépnek versenybe az aktív molekulákkal.
Génsodródás - a gének (allélok) gyakoriságának változása kis populációkban a párok véletlenszerű kombinációja miatt a szaporodás során. Egy populáció genotípusos szerkezetének genetikai sodródásból adódó dinamikájának jellegzetes vonása a homozigotizáció növekedése, amely a populáció méretének csökkenésével növekszik. Tekintsük egy hipotetikus populáció viselkedését, amelyben a Hardy-Weinberg-formula szerinti genotípusok 1AA + 2Aa + 1aa arányban jelennek meg. Ha ebből a populációból csak két egyed vesz részt a keresztezésben, akkor annak a valószínűsége, hogy egy vagy másik genotípus szüleit egy párban kombinálják, a következő lesz:

A táblázatból következik, hogy ennek a hipotetikus populációnak a homozigóta állapotába való átmenet valószínűsége: AA × AA = 1/16; aa × aa = 1/16, ami 1/16 + 1/16 = 2/16 = 1/8. Ez azt jelenti, hogy bármely allél véletlen elvesztésének valószínűsége 1 generáció alatt 1/8, azaz átlagosan 8 generáció alatt a populáció AA vagy aa lesz. A genetikai sodródás szerepe a populáció genotípusának megváltoztatásában gyorsan csökken a populáció effektív méretének növekedésével.
50 egyedből álló populáció mellett a genetikai sodródás miatt generációnkénti heterozigóták gyakorisága 0,01, 500 egyedből álló populáció esetén 0,001, azaz 0,1%-kal csökkenhet. A heterozigóták gyakorisága egy populációban a genetikai sodródás következtében egy generáció alatt csökken a K = 1/2n képlettel kifejezett mennyiséggel, ahol K a heterozigóták gyakoriságának csökkenésének aránya, n pedig a populáció tényleges mérete.
Megkettőzés – egy gén vagy egy génszakasz megkettőzése. Lásd Mutation, Crossing over.
Az eugenika az emberi örökletes egészség doktrínája és annak javításának módjai.
Zigóta - a különböző nemű ivarsejtek fúziója eredményeként létrejövő sejt; megtermékenyített petesejt.
A Zygoten az I. próféta felosztásának egyik szakasza.
A zigóta DNS az eukarióta DNS része, amely részt vesz a heteroduplex, azaz egy hibrid kettős hélix kialakulásában, melynek fonalai a meiózis során különböző kromoszómákhoz tartoznak, ami késlelteti replikációját addig, amíg a zigotánban szinaptonemális komplex nem jelenik meg.
A zigóta indukció egy fág indukálása, amelyet egy prófégot hordozó donor, például λ lizogén baktérium kromoszómafragmensének behatolása okoz a nem lizogén befogadó sejtbe, amely indukálódik és vegetatív állapotba kerül, ami a zigóta líziséhez. Ennek eredményeként az ilyen keresztezésekben csak azok a markerek jelenhetnek meg a rekombinánsokban, amelyeket a prófég előtt továbbítottak.
Az idiogram egy kariotípus sematikus általánosítása az egyes kromoszómák és részeik közötti átlagos mennyiségi összefüggések megfigyelésével.
Az izoakceptor tRNS-ek különböző tRNS-ek, amelyek ugyanazt az aminosavat hordozzák a fehérjeszintézis során, különböző kodonokat ismernek fel az mRNS-en, és ezért eltérő antikodonokkal rendelkeznek. Például két izoakceptor leucin tRNS. egy nyúl májából származó leucint tartalmaznak a szintetizált polipeptidlánc különböző helyein, mivel az egyikük felismeri az mRNS-t. a CUU hármas, a másik pedig a CUU hármas. Ezen tRNS-ek aránya határozza meg a fehérjeszintézis lefolyását.
Az izoleucin egy esszenciális aminosav, amely szinte minden fehérjében megtalálható.
Izoláció - ugyanazon faj egyedei közötti szabad keresztezés kizárása vagy nehézsége, ami fajon belüli csoportok és új fajok izolálásához vezet.
Az izomerázok az enzimek egy osztálya, amelyek katalizálják a szerves vegyületek intramolekuláris átrendeződési reakcióit, beleértve az izomerek interkonverzióját.
Izoenzimek - azonos vagy hasonló funkciójú enzimek, amelyeket ugyanazon kromoszómakészlet (haploid) különböző lókuszai (génjei) kódolnak. Az ilyen lókuszok a duplikáció és az azt követő változások eredményeként jönnek létre. Tehát egy egyedben egy enzim több változata is szintetizálható. Az izoenzimek megtalálhatók az állatok, növények szöveteiben és mikroorganizmusokban is. Fontos szerepet játszanak az enzimaktivitás szabályozásában, valamint a fejlődési folyamatokban. Az izoenzimkészlet az állatok és növények különböző szövetei és szervezetei között genetikailag meghatározott (mennyiségi és minőségi) különbségek eredményeként jön létre, és gyakran szigorúan specifikus. Egy bizonyos izoenzim jelenlétét vagy hiányát széles körben használják genetikai markerként annak meghatározására, hogy egy egyed egy bizonyos csoporthoz tartozik-e, és egy fehérje izoenzimének gyakoriságának elemzését használják a populációk határainak meghatározására.
Az immobilizált enzimek mesterségesen előállított enzimkészítmények, amelyek molekulái kovalensen kötődnek egy polimer hordozóhoz, aminek következtében a denaturáló hatásokkal szembeni ellenállásuk jelentősen megnő.
Immunogenetika - szakasz orvosi genetika, a humán immunológiai rendszerek, vércsoportok genetikai meghatározottságának vizsgálata.
Az immunglobulinok összetett fehérjék (glikoproteinek), amelyek specifikusan kötődnek idegen anyagokhoz - antigénekhez.
A beltenyésztés közeli rokon egyedek keresztezése.
Inverzió - a kromoszóma-átrendeződés egy fajtája, amely a genetikai anyag egy részének 180 ° -os megfordításából áll.
Inhibitorok - különböző kémiai természetű anyagok, amelyek elnyomják az egyes enzimek vagy enzimrendszerek katalitikus aktivitását.
Indikátor törzs – más baktériumtörzsek vagy bakteriofágok azonosítására (izolálására) használják.
Indukció - a baktérium- és élesztősejtek azon képessége, hogy bizonyos enzimeket csak megfelelő szubsztrátok jelenlétében szintetizáljanak a környezetben, transzkripcióváltás az induktor és a szabályozó fehérje kölcsönhatása következtében; a profág kivágása a gazdasejt genomjából a represszor fehérje inaktiválása miatt, ami a fejlődés litikus ciklusának kezdetéhez vezet.
Indukálható enzimek - enzimek, amelyek szintézisének sebessége a szervezet körülményeitől függően változik. A fehérjeszintézis beindítása. Lásd: Közvetítés.
Initiációs kodon - a legtöbb vagy az összes polipeptid lánc szintézise Escherichia coliban, amely a formil-metionin amino-terminálisba történő beépülésével kezdődik az AUG iniciációs kodon jelenléte miatt, amelyet a Met-tRNS met felismer. Lásd: Közvetítés.
beillesztési szegmensek. Lásd az IS elemeket.
Inszerció - egy mozgó genetikai elem beillesztése egy kromoszómán vagy plazmidon valamilyen új helyre; gyakran inszerciós génmutációk kialakulásához vezet.
Interkinézis - a meiózis első és második felosztása közötti időszak.
Interszex - a szexuális jellemzők köztes megnyilvánulásával rendelkező egyének.
Az interperszió az eukarióták kromoszómális DNS-ében lévő, különböző fokú ismétlődésű nukleotidszekvenciák váltakozása.
Az interfázis a meiotikus ciklusnak az egyik meiózis vége és a következő kezdete közötti szakasza. Három szakaszból áll: preszintetikus, DNS-szintézis és posztszintetikus.
Interferencia. Lásd Crossover.
Az intron egy meghatározott nukleotidszekvencia inszerciója egy génen belül, amely nem hordoz genetikai információt. Az eukarióták génjeiben található. Az intronok hossza nagyon változó. Ezek teljes hossza gyakran nagyobb, mint az információt hordozó gén többi részének (exon) hossza. A nitronok és az exonok közötti határ a nukleotidok bizonyos kombinációja mentén fut (TT - az egyik végétől, GC - a másik végétől). Az érett mRNS-molekula megjelenése az intronok eltávolítása után az egyes szakaszok keresztkötésének eredménye. Ezt a folyamatot splicingnek nevezik. Bizonyíték van arra, hogy az intronok nem a gén nem funkcionális részei. Például az élesztőben a mitokondriális citokrómot szabályozó génben az intronok más fehérjéket kódolnak, amelyek a citokróm mRNS érési folyamatában működnek ("önkiszolgálás"). Az intronok jelenléte fontos a genetikai rekombinációs folyamatok szempontjából, ami a káros mutációk semlegesítéséhez és új gének kialakulásához vezethet.
A mesterséges szelekció egy adott fajhoz, fajtához vagy fajtához tartozó állatok és növények gazdaságilag legértékesebb egyedeinek kiválasztása annak érdekében, hogy azokból kívánatos tulajdonságokkal rendelkező utódokat kapjon.
IS-elemek - transzponáló elemek (TE), amelyek mérete 200-5 × 10 3 pár nukleotid, és csak a transzpozícióhoz kapcsolódó géneket tartalmazzák. A kromoszóma különböző helyeire integrálódva az IS-elemek nemcsak a közeli géneket inaktiválhatják, amelyek fenotípusosan látható vagy letális mutációként fejeződhetnek ki, hanem kapcsolók szerepét is betölthetik, megállítva vagy lehetővé téve ezeknek a géneknek a transzkripcióját, attól függően, hogy a maga az IS transzkripció iránya egybeesik vagy sem.-elem és bakteriális operon, az IS 2 -elem így szabályozhatja a galaktóz operon munkáját.
Kariotípus - a kromoszómakészlet jellemzőinek halmaza (szám, méret, forma), amely egy adott kromoszómatípusra jellemző.
Komplementációs térkép - az allélmutációk közötti kapcsolat lineáris vagy körkörös, nem átfedő (komplementáció hiányában) szegmensei formájában lévő kép, amely tükrözi a tercier szerkezetben elhelyezett polipeptidláncok kölcsönhatását. A fehérje tercier és kvaterner szerkezetének genetikai elemzésének módszere az interallélikus komplementáció vizsgálatán alapul. Az interallél komplementációs teszt a fehérjék evolúciós divergenciájának vizsgálatára használható. Például az E. coli és a Serratla marcescens alkalikus foszfatázai ugyanazokat a reakciókat hajtják végre, de eltérő aminosav-összetételűek. Komplementáció van közöttük, ami mindkét faj alkalikus foszfatáz alegységei közötti kölcsönhatási központok evolúciós konzervativizmusát jelzi. Hasonlóképpen, aktív triptofán-szintetáz nyerhető az E. coli heterológ calamus alegységeiből és az enterobaktériumok más képviselőiből.
Mapping - a kromoszómán lévő gének közötti lokalizáció (sorrend és kölcsönös távolság) meghatározása (génen belüli mutációk). Létezik heteroduplex és restrikciós leképezés. Heteroduplex térképezés - térképezés két különböző, de közeli genom DNS-szegmensének hibridizációjával. A renaturált DNS-molekulák között előfordulhatnak olyan szerkezetek, amelyekben a DNS egyes szakaszai a komplementaritás hiánya miatt nem kapcsolódnak kettős szálú molekulává. Ezeket a területeket elektronmikroszkóp alatt határozzuk meg. Meg lehet állapítani hosszukat és helyzetüket a genom többi részéhez képest. Heteroduplex térképezés akkor is elvégezhető, ha a DNS-t a megfelelő mRNS-sel hibridizáljuk. A feltérképezést az egyszálú hurkok lokalizációja szerint végezzük. Restrikciós térképezés - egyes élőlények genomja (vírusok, mitokondriumok, kloroplasztiszok, nagyobb genomok részei) enzimatikusan külön fragmentumokra hasítható. A kis fragmentumok összehasonlításakor a teljes genom komponenseinek sorrendje megállapítható.
Az operonok kaszkádszabályozása - a transzkripció váltása egyik szerkezeti génről a másikra a vírusok és prokarióták életciklusa során.
A katabolizmus egy élő szervezetben végbemenő enzimreakciók összessége, amelyek az élelmiszerrel szállított vagy a szervezetben tárolt összetett szerves anyagok - fehérjék, nukleinsavak, zsírok, szénhidrátok - lebontását célozzák.
Katabolit represszió - az adaptív enzimek szintézisének gátlása baktériumtenyészetekben glükóz jelenlétében, az úgynevezett glükózhatás. A hatásért felelős inhibitor egy bomlástermék, azaz a glükóz katabolizmus. Glükóz hiányában az adenilát-cikláz enzim hatására az ATP-ből ciklikus 3'-5'-AMP vagy cAMP képződik, amely a BAC fehérjével kombinálva a promoterhez kötődik, és lehetővé teszi az RNS polimeráz transzkripciójának megindítását. . Glükóz jelenlétében az adenilát-cikláz aktivitása elnyomódik, ami a cAMP koncentrációjának csökkenéséhez vezet, és megakadályozza az adaptív enzimek transzkripcióját. A glükóz hatás a laktózt, maltózt, arabinózt, xilózt, galaktózt tartalmazó tápközegben is megfigyelhető. Fruktózon és mannózon nem figyelhető meg.
A katénok nukleinsavak láncok formájában lévő szerkezetei. Például számos vírus (SV40, FX174) és mitokondriumok körkörös DNS-molekulái ugyanabban a láncban kapcsolódnak egymáshoz.
Klaszter - funkcionálisan kapcsolódó gének egy sorban elrendezve.
A sejtmanipuláció egy új típusú sejtek létrehozásának módszere azok tenyésztése, hibridizációja és rekonstrukciója alapján.
Klón – sejtek vagy egyedek halmaza, amelyek ivartalan szaporodás útján egy közös őstől származnak.
A molekuláris klónozás a rekombináns DNS-molekulák kimutatásának módszere olyan tápanyag-agar-sejteken történő szűréssel és tenyésztéssel, amelyekbe transzformációval ilyen DNS-t juttattak be. Baktériumok esetében minden sejt kolóniát képez, ami egy klón, amelynek minden sejtje azonos rekombináns DNS molekulákat tartalmaz.
Kodominancia - mindkét allél részvétele a tulajdonság meghatározásában egy heterozigóta egyedben (klasszikus példa egy bizonyos vércsoport MM, MN, NN alléljainak kölcsönhatása).
Kodon - hármas; a genetikai kód diszkrét egysége; három szekvenciából álló hírvivő RNS darab.
Co-incidens – a megfigyelt átkelések (keresztezések) számának aránya az elméletileg várthoz képest. Lásd Crossover.
A gén és az általa kódolt fehérje kolinearitása a fehérjében lévő aminosav-maradékok DNS-nukleotidok váltakozása általi elrendeződésének feltétele.
Colicines. Lásd: Bakteriocin.
Kombinatív variabilitás - variabilitás, amely rekombinációk kialakulásán alapul, vagyis olyan génkombinációk, amelyekkel a szülők nem rendelkeztek.
Vegyület - egy heterozigóta több allél sorozatából származó két mutáns allél vegyületet képez (w a /w ch).
A gének dózisainak kompenzálása - az X kromoszómán elhelyezkedő géncsoportok aktivitásának szabályozása. A nőstény emlősök ontogenezisében mindkét X kromoszóma génje csak az embriogenezis korai szakaszában aktív, amikor eldől, hogy a szervezet későbbi differenciálódása inkább a nőstény, mint a hím nem felé irányul. Később a nőknél az X kromoszómák egyike heterokromatizálódik, és a benne lokalizált gének átírása megszűnik. Ennek eredményeként a homogametikus nemben, akárcsak a heterogametikus nemben, csak egy, az X kromoszómán fekvő génkészlet nyilvánul meg fenotípusosan. Az elsődleges és másodlagos nemi jellemzők későbbi fejlődését a nemi hormonok határozzák meg. Két X-kromoszóma aktivitása nemcsak a nemben, hanem más életjelekben is nagy különbségek kialakulásához vezetne. Meg kell jegyezni, hogy az X kromoszómával kapcsolatban a heterogametikus egyedek monoszómák. Ezzel mindkét nemnél azonos a nemi hormonoktól független jelek kialakulása. A Drosophilában a hímek és nők X kromoszómájában lokalizált gének aktivitásának egyenletét azzal érik el, hogy a férfiaknál az X kromoszóma génjeinek aktivitása kétszerese. A hímek X-kromoszómájának génjeinek aktiválása egy aktiváló faktor (valószínűleg fehérje jellegű) jelenléte miatt történik.
Az illetékes sejtek olyan sejtek, amelyek képesek az átalakulásra. Lásd: Átalakítás.
beavatási komplexus. Lásd: Közvetítés.
A komplementaritás az egymást kiegészítő gének jelenléte, amelyek kombinálva egy új (vad) tulajdonság megjelenését határozzák meg. A komplementer interakcióban a hasítás 9:7, 9:3:4, 9:3:3:1 lehet. A gének primer termékeik szintjén történő kölcsönhatására példa a komplementáció, amely egyes kvaterner szerkezetű, azaz több azonos vagy különböző polipeptidláncból álló fehérje szintézise során nyilvánul meg (az alkalikus foszfatáznak 2 egyforma lánca van, hemoglobin - 4 kétféle lánc). Két különböző gének mutáns szülőjének hibridje képes normálisan működő fehérjét szintetizálni (AAbb × aaBB → AaBb). Nemcsak különböző gének komplementálhatják, hanem ugyanazon gén különböző alléljai is. Ilyen interallél komplementáció akkor következik be, amikor egy gén allélmutációi különböző módon megváltoztatják az aminosavak sorrendjét a polipeptidláncban, ami torzítja annak másodlagos és harmadlagos szerkezetét, megfosztva az enzim aktivitásától. Azon heterozigótákban, amelyek genomjában a gén különböző alléljei vannak, a megváltozott láncok térbeli konfigurációjának normalizálódása figyelhető meg, és egy több alegységből álló hibrid molekula teljes vagy részleges aktivitású lehet. Lásd a kiegészítési térképet.
Komplementer gének - két vagy több nem allél gén, amelyek fenotípusos expressziója szükséges a szervezet egy tulajdonságának kifejeződéséhez.
Komplementer DNS (cDNS) – in vitro reverz transzkriptáz (vagy DNS-polimeráz) segítségével szintetizált mRNS-másolatok, amelyek megfelelnek egy specifikus génnek intronok nélkül.
A komplementáció egy vad vagy közeli fenotípus helyreállítása, amikor két különböző vagy hasonló fenotípusos megnyilvánulású recesszív mutációt kombinálnak egy sejtben.
A konvergencia az élőlények különböző csoportjaiban hasonló tulajdonságok független fejlődése hasonló környezeti feltételekhez.
A génkonverzió olyan keresztezés, amely egy heterozigóta gén közelében ment végbe, és amelynek alléljeinek felhasadása megsérti a reciprocitást. A neurospórák génkonverziója során a 6A:2a, 2A:6a, 5a:3a hasítás figyelhető meg az ascusban a normál 4A:4a helyett.
A konkordancia a vizsgált tulajdonság megnyilvánulása egy pár mindkét ikerpárjában.
Konstitutív szintézis - enzimek szintézise induktor hiányában. Amikor a lac operonban az i - mutáció konstitutív szintézishez vezet, a megváltozott szabályozó fehérje nem kötődik az operátorhoz, és amikor az O c mutáció bekövetkezik, a normál szabályozó fehérje (represszor) nem kötődik a mutáns operátorhoz. Ilyen esetekben a szerkezeti gének elérhetővé válnak az RNS-polimeráz számára, és átíródnak. A sejt által folyamatosan szintetizált enzimeket, függetlenül attól, hogy szubsztrát vagy induktor van a környezetben, konstitutívnak nevezzük.
A baktériumok konjugálása a genetikai anyag cseréjének egyik módja, amelynek során egyirányú átvitele történik a donortól ("férfi" sejt) a recipiensig ("női" sejt).
A kromoszómakonjugáció a homológ kromoszómák páronkénti átmeneti megközelítése, amelyben lehetséges a homológ régióik cseréje - átkelés.
Terminális redundancia - bázisok vagy gének ismétlődő szekvenciáinak jelenléte a kromoszóma végén. fágokban figyelhető meg.
Az enzimek összehangolt elnyomása - egy enzim szintézisének leállítása az általa katalizált reakciótermék jelenlétében. Azt találták, hogy a trp A és trp B gének által meghatározott enzim, az E. coli triptofán-szintáz szintézise gátolt triptofán jelenlétében. A jelenség biológiai jelentése nyilvánvaló: gazdaságtalan lenne, ha egy sejt olyan enzimeket szintetizálna, amelyek biztosítják a triptofán szintézisét, mivel ezt az aminosavat elegendő mennyiségben tartalmazza a táptalaj. A triptofán koordináció szabályozza mind az öt szomszédos gén trp A, B, C, D, E aktivitását. Ez azt jelenti, hogy a triptofán szintetáz (trp A, trp B), IGF szintetáz (trp D), foszforibozil antranilát transzferáz (trp) intracelluláris tartalma. A C) és az antranilát-szintáz (trp E) a triptofán-koncentráció változásával azonos mértékben változik. Represszálható operon esetén a represszor inaktív állapotban van triptofán hiányában, és aktívvá válik, amiben triptofán jelenlétében egy operátorhoz tud kötődni.
A koenzimek nem fehérje szerves vegyületek, amelyek egyes enzimek aktív központjának részét képezik.
Co-evolúció - különböző fajokhoz tartozó organizmusok evolúciós kölcsönhatásai, amelyek nem cserélnek genetikai információt, de biológiailag szorosan összefüggenek.
örökölhetőségi együttható. Lásd: Örökölhetőség.
A keresztirányú (cross-cross) öröklődés a nemhez kötött vonások apáról leányaira és anyáról fiúkra történő átvitelének egyik fajtája.
A rejtélyes mutáns egy lac Z Y + mutáns, amelyből hiányzik a galaktozid-permeáz, ezért nem szintetizál β-galaktozidázt. A laktóz hidrolizálásának képessége ebben a mutánsban csak sejtkivonatokban található meg.
A kriptafágok (rejtett fágok) a profág genom része, amely a bakteriális kromoszómában marad a hibás transzdukáló fág kialakulása után. A kriptafágok képesek rekombinációra más mutáns homológ fágokkal.
A kriptoméria a génkölcsönhatások (recesszív episztázis) egyik fajtája.
A meiózis során a homológ kromoszómák metszeteinek kölcsönös cseréje, ami új génkombinációk és ezt követően rekombináns egyedek megjelenéséhez vezet, a gének közötti távolságtól függ, és a kromoszóma-térképezés mértékeként szolgál.
Egyenlőtlen keresztezés – a csere megfelelőségének megsértése az átkelés során különböző hosszúságú kromoszóma szakaszok cseréjéhez vezet, ami a duplikációk oka.
bal operátor. Lásd a kezelőt.
Liázok - az enzimek egy osztálya, amelyek katalizálják bizonyos atomcsoportok szubsztrátjainak nem hidrolitikus hasadási reakcióit a képződéssel kettős kötések, valamint atomok és atomcsoportok kettős kötésekhez való addíciós reakciói.
A lektinek olyan növényi fehérjék, amelyek a sejtfelszín szénhidrát komponenseihez való szelektív kötődés eredményeként aggregálják az emlős sejteket.
A ligázok az enzimek egy osztálya, amelyek a nukleozid-trifoszfátok hidrolízisének kapcsolt reakciójának energiája miatt két különböző molekula egymáshoz kapcsolódási reakcióit katalizálják. A DNS javítására, replikációjára és rekombinációjára használják. Katalizálja a foszfodiészter kötések szintézisét a kétszálú DNS szomszédos 3'-hidroxil- és 5'-foszfát-végei között. Például egy E. coli sejt legfeljebb 400 ligáz molekulát tartalmaz. A DNS-ligázokat eukarióta sejtekből is izolálják. Az emlőssejtekben kétféle ligáz létezik, amelyek szerológiailag különböznek egymástól: a ligáz I elsősorban a citoplazmában, a ligáz II pedig a sejtmagban és a mitokondriumokban lokalizálódik.
A vezető szekvencia az mRNS 5' végén lévő, az iniciációs kodont megelőző nem transzlált régiója.
Leader RNS - az RNS egy része (90%), amelynek transzkripciója a triptofán koncentráció gyors ingadozása során az attenuátor régióban (például a trp-operonban) véget ér. Mély triptofán-éhezés körülményei között a csillapítóban a terminációk aránya nullára csökken, és a beindított transzkripciók a szerkezeti cisztronok zónájába kerülnek, biztosítva ezzel a triptofán szintézisét. Lásd Csillapító.
Vezető DNS-szál (leading) - újonnan szintetizált DNS-szál, amelynek iránya (5'-3') egybeesik a replikációs villa mozgási irányával. A második, újonnan szintetizált szálat, amely komplementer az első szálhoz, laggingnek nevezzük.
A HFT-lizátum olyan lizátum, amelyet kétszeresen lizogén baktériumok ultraibolya fénnyel történő indukciójának eredményeként nyernek, és amely k-gal-hibás és λ-ép profágokat tartalmaz. Magas átviteli képességgel rendelkezik.
A lizogén egy baktériumsejt állapota, amelyben egy vagy több bakteriofág található a kromoszómáján.
A lizogén konverzió új tulajdonságok (tulajdonságok) megszerzése egy baktériumsejt által a lizogenizáció során. A lizogén sejt rezisztenssé válik a homológ fággal szemben; a lizogén K12 (λ) törzsek nem képesek támogatni a T-even fágok rII mutánsainak növekedését; a PI fágra lizogén sejtek módosítják a λ fág DNS-ét. Ezek a módosítások megvédik a λ fág DNS-t a PI restrikciós nukleáz általi lebomlástól, amely lebontja a normál, módosítatlan λ fág DNS-t. A PI-fágra lizogén sejtben túlélő, módosított DNS-sel rendelkező ritka λ-részecskék leszármazottait a lizogén KPI-törzsek már nem korlátozzák, és elszaporodnak rajtuk. Lásd: Gazda által vezérelt módosítások.
A lizozim a hidroláz osztályba tartozó enzim; katalizálja az aminocukor-maradékok közötti β-1,4-glikozidos kötések hidrolízisét. N-acetil-glükózamin és N-acetilmuraminsav a mureinok poliszacharid láncaiban, ami a baktériumsejt membrán tönkremeneteléhez vezet.
Linkerek. Lásd a ragadós végeket.
Tiszta vonalak - genotipikusan homogén organizmusok összessége, amelyek növények önbeporzásából vagy állatok hosszú távú beltenyésztéséből származnak.
A ragadós végek átfedő végrégiók, amelyek felelősek a λ fág lineáris kromoszómájának körkörössé való átalakulásáért. A ragadós végek hossza eléri a 12 nukleotidot. A λ fág A génjében kódolt fágspecifikus nukleáz (endonukleáz) hatására jönnek létre, és a gazdasejtben csak a fág vegetatív szaporodása során képződnek. A génsebészetben úgynevezett linkereket használnak - rövid szintetikus oligonukleotidokat. A polilinkereket, amelyek számos restrikciós endonukleáz felismerési helyei, gyakran beépítenek vektorokba.
Lokusz - egy adott mutáció helye a kromoszóma genetikai vagy citológiai térképén. Ez a fogalom relatív, és két mutációt úgy tekintünk, hogy ugyanabban a lókuszban helyezkednek el mindaddig, amíg meg nem állapítják közöttük az átkelés lehetőségét. Gyakrabban használják a kromoszóma nagy régióira, amelyek egy teljes gént tartalmaznak.
A gének nagyítása - az rRNS gének lehasítása a kromoszómáról körkörös molekulák formájában, replikációjuk a kromoszómába való későbbi integrációval. Petesejtekben, embrionális sejtekben figyelhető meg a fejlődés korai szakaszában. Megnövekedett számú gén kerül át az ivarsejteken keresztül a következő generációkba. Ez az rRNS-gének számának kompenzációs növekedése válaszul az egyenlőtlen keresztezés eredményeként bekövetkező elvesztésükre. Drosophilában látható.
Anyai öröklődés - extrakromoszómális (citoplazmatikus) faktorok által szabályozott öröklődés, amely fenotípusos különbségekhez vezet az azonos genotípusú egyének között.
Interline hibridek - beltenyésztett vonalak keresztezéséből nyert hibridek. Az interlineáris hibrideket, például a kukoricát az első generációban értékelik a heterózis hatásának szempontjából, kiválasztják a legjobb kombinációt adó vonalakat, majd nagy léptékben szaporítják hibrid magvak előállítására. A hibrid magok átvételekor a kezdeti vonalakat sorba vetik, váltakozva az anyai és apai formák között. A köztük lévő beporzás biztosítására citoplazmatikus hímsterilitást (CMS) használnak. A kettős sorközi hibrideket széles körben használják. Két egyszerű, heterózist mutató hibrid keresztezésével nyerik őket. Az ilyen kettős hibridek gyakran heterózist mutatnak, és négy különböző fajtából származó négy beltenyésztett vonal használatán alapulnak: (A × B) × (C × D).
A meiózis a sejtosztódás egy speciális módszere, melynek eredményeként a kromoszómák száma csökken (csökken), és a sejtek diploid állapotból haploid állapotba kerülnek. A növényekben a meiózis a sporofitákban fordul elő mikro- és makrospórák képződésével, állatokban - a nőstényekben a petesejtekben és a hímekben a spermatocitákban az úgynevezett érés során. A diploid és a haploid fázisok aránya szerint a szervezet életciklusában háromféle meiózist különböztetnek meg: 1) kezdeti, vagy zigóta (közvetlenül a megtermékenyítés után a zigóta első osztódásaival, algákban és protozoonokban fordul elő); 2) köztes vagy spóra (a legtöbb növényben a spóraképződés időszakában fordul elő a sporofita és a gametofita szakaszai között); 3) végső, vagy gametikus (minden többsejtű állatra, egyes protozoákra és alacsonyabb rendű növényekre, például barna algákra jellemző).
A melaninok fekete, barna vagy sárga pigmentek. A melanin molekulák összetett komplexek, amelyeket tirozinszármazékok és fehérjék polimerei alkotnak.
A mendelizmus a szervezet jellemzőinek öröklődési törvényeinek doktrínája.
A merodiploid egy részleges diploid.
A merozigóta olyan részleges zigóta, amely a baktériumokban a transzformáció, transzdukció és szexdukció során fordul elő, amikor a donor sejtnek csak egy DNS-fragmentuma, amely egy vagy több gént tartalmaz, kerül a befogadó sejtbe. Ha a kromoszómával konjugált bejutott hely (exogén) allél összetételében eltér a bakteriális kromoszóma megfelelő szegmensétől (endogén), akkor részleges heterozigóta képződik, amelyet heterogénnek is neveznek.
A migráló vagy mobil genetikai elemek (MGE) a genetikai anyag azon részei, amelyek egyetlen sejten belül képesek mozogni a genomon belül. A mutációk és variációk a különböző eredetű MGE-k mozgásaihoz kapcsolódnak (baktériumokban IS-elemek és transzpozonok, különböző állatokban mobil diszpergált gének, növényekben olyan elemek, mint aktivátor és disszociátor). A variációk például abban fejeződnek ki, hogy a természetes Drosophila populációkban az egyes egyedek eltérnek az MGE-k elhelyezkedésében és számában. Az MGE vírusok, különösen a Drosophila szén-dioxiddal szembeni érzékenységét meghatározó szigmavírus inszerciója ugyanazon mutációk szinkron "kitörésének" jelenségével függ össze a különböző földrajzi területeken élő, azonos fajhoz tartozó szervezetekben.
Mikroszómák - sejthomogenizátumok differenciális centrifugálásával nyert szubcelluláris frakció.
Minisejtek – az E. coli mutánsai, hibásak a szaporodási képességükben, nem tartalmaznak DNS-t és nem képesek osztódni. A normál sejt térfogatának körülbelül 10%-át teszik ki.
Kisebb alapok. Lásd ritka alapokat.
Mínusz lánc. Lásd plusz-mínusz láncot.
A mitózis (közvetett osztódás) az eukarióta sejtek osztódásának fő módja. A mitózis biológiai jelentősége a reduplikált kromoszómák szigorúan azonos eloszlásában rejlik a leánysejtek között, ami biztosítja a genetikailag ekvivalens sejtek kialakulását és a folytonosságot számos sejtgenerációban.
Egy gén többszörös működése. Lásd a gén pleiotróp hatása.
A fertőzés többszöröse az egy baktériumsejten adszorbeált fágrészecskék száma.
A mobil gének szerkezetileg és genetikailag különálló DNS-fragmensek, amelyek a sejtgenom körül mozoghatnak.
A módosítások egy szervezet jellemzőiben a környezeti tényezők hatására bekövetkező változások, de nem befolyásolják a genotípust. A módosítások a szervezet egyértelmű reakcióit jelentik a környezet hatására. Nem öröklődnek, és a szervezet élete során fennmaradnak. A tulajdonság változásai bizonyos határok között ingadoznak, ami genotípustól függ, és ezt reakciónormának nevezzük. Mert különböző jelek a reakciósebesség más.
A módosulások vizsgálata a genotípus és a környezet kölcsönhatásának relatív szerepének és természetének tisztázásához, az evolúció törvényeinek megértéséhez szükséges, hiszen a természetes szelekció a fenotípus szintjén hat, azaz mutációkkal és módosulásokkal egyaránt operál. A módosítások példái közé tartozik a klorofillszemcsék számának növekedése, amikor a levelek egy részét eltávolítják a szár és a levéldugványok asszimiláló szöveteiben, a len, a kínai kankalin és a lepkék szirmai színének változása a hőmérséklet függvényében, a növények megvastagodott és gyér termésekben. A módosítások akkor alkalmazkodóak (adekvátak), ha azokat a környezet szokásos változásai okozzák, amelyeknek egy adott faj egyedei ki voltak téve a múltbeli evolúciós története során. Ha a szervezet olyan szokatlan körülmények közé kerül, amelyekkel még nem találkoztak ezt a fajt, akkor adaptív jelentőséggel nem rendelkező módosulások léphetnek fel (például a nyílhegy gyenge fényben fejlődő légi levelei ugyanolyan szalagszerű alakúak, mint a víz alattiak). Léteznek morfózisnak nevezett káros módosulások is – a faj számára extrém vagy szokatlan környezeti tényezők által okozott nem örökletes változások, különösen a bórtöbblet egyes növényekben klorózishoz vezet, a lítium-klorid jelenlétében fejlődő halivadékban csak az egyik szem kialakult (ciklopizmus) . Egyes morfózisok hasonlíthatnak bizonyos gének fenotípusos hatására. Fenokópiáknak nevezik őket, például a Drosophila pupae hőmérsékleti sokk hatására felfelé kunkorodó és széttárt szárnyú legyeket kapnak, a vg vonalnál megnagyobbodott szárnyakkal.
A tulajdonos által irányított módosítások. - Az E. coli K-12-n tenyésztett I. fág legtöbb részecskéje (nevezzük őket λ K-nek) nem képes elszaporodni a PI-profágot hordozó E. coli K-12 (PI) lizogén baktériumon. A K-12-n (PI) elszaporodott ritka λ-részecskék leszármazottai (nevezzük őket λ PI-nek) mindkét törzsen felnőhetnek, azaz nem korlátozódnak, DNS-üket nem bontja le a PI restrikciós enzim. Az ilyen rezisztencia a fág DNS módosulásának következménye, vagyis az 5-metil-citozin és a 6-metil-aminopurin kisebb bázisainak megjelenése a citozin és az adenin metilezése következtében, a metilcsoportot átvivő bakteriális metiltranszferázok hatására. Az S-adenozil-metionint két közös bázisra (adeninre és citozinra) hozzuk létre. Lásd lizogén konverzió.
Transláció utáni módosítások. - A riboszómán képződött fehérje gyakran befejezetlen marad, és ezt követően enzimatikus módosuláson megy keresztül. Az N-terminális fMet vagy Met lehasad, a szekréciós fehérjék elvesztik "jelszekvenciájukat", és szénhidrátokból fedezik. Egyes enzimek (pepszin, zimotripszin, tripszin, inzulin) hosszabb prekurzorként képződnek, mint késztermék, amely a sejt önvédelme ezen enzimek aktivitása ellen. Az alloszterikus fehérjék aktivitása megváltozik, ha szubsztrátot (effektort) kötnek hozzájuk. A végső tercier és kvaterner struktúrák kialakulása diszulfidhidak képződésével, és néha ritka aminosavak képződésével történik, amelyek nem rendelkeznek saját kodonnal (hidroxiprolin a prolinból).
Mozaikok - különböző genotípusú sejtekből álló organizmusok; mutációk vagy szomatikus keresztezés következtében alakulnak ki.
Csendes mutációk - olyan mutációk, amelyek nem nyilvánulnak meg fenotípusosan, és a mutációk gyakoriságának látható csökkenéséhez vezetnek Tone S - Tone r (érzékenység - rezisztencia a TI fággal szemben). Ezek a TI fág receptorainak szerkezetét meghatározó fehérje változásain alapulnak, amelyek nem elegendőek a Ton r fenotípus meghatározásához.
A mutagén olyan fizikai vagy kémiai anyag, amely növeli a mutációk gyakoriságát.
A mutagenezis a mutációk előfordulásának folyamata.
A mutáns olyan sejt vagy egyedi organizmus, amelyet egy mutáció okozta változás jellemez.
A mutáció olyan genetikai változás, amely a genetikai anyag alapvető tulajdonságainak minőségileg új megnyilvánulásához vezet.
A generatív mutáció olyan mutáció, amely a generatív sejtekben fordul elő, és öröklődik.
Genetikai mutáció - olyan mutáció, amelyben az egyes gének szerkezete átrendeződik.
A mutáció genomikus. Lásd: poliploidia.
A missense mutáció a kodonok nukleotid összetételének megsértése, amelyben a megváltozott kód meghatározza a rossz aminosav beépülését a szintetizált fehérjébe.
A nonszensz mutáció olyan kodonváltás, amelyben az új kodon egyáltalán nem határozza meg egyetlen aminosav beépülését sem. A nonszensz mutációk véget érnek, és a következő kodonok megjelenéséhez vezetnek: UAG - borostyán, UAA - okker, UGA - opál.
A reverz mutáció vagy reverzió olyan mutáció, amely a vad fenotípus helyreállításához vezet.
Frameshift mutáció - inszerciók (inszerciók) vagy deléciók (deléciók), amelyek a transzkripció és ennek megfelelően a szintetizált polipeptidek szerkezetének torzulásához vezetnek.
A szomatikus mutációk olyan mutációk, amelyek szomatikus sejtekben fordulnak elő, és nem öröklődnek.
Mu-mutagenezis - mutagenezis a Mu bakteriofág hatása alatt.
Az öröklődés egy élőlény örökletes jeleinek és tulajdonságainak hajlamainak átvitelének folyamata a szaporodási folyamat során.
Az öröklődés a sejt- és testszerkezet olyan tulajdonsága, amely biztosítja a generációk közötti anyagi és funkcionális folytonosságot.
Az örökletes betegségek génmutációval összefüggő kóros állapotok, amelyek generációról generációra terjednek.
Örökölhetőség – egy adott tulajdonság genetikai szabályozásának mértéke, azaz a genetikai variabilitás és a fenotípus aránya.
Negatív szintézis indukció - indukció, amely abból áll, hogy a szabályozó fehérje (represszor) induktor (szubsztrát) jelenlétében nem kötődik az operátorhoz (az allosztérikus hatás miatt), így lehetővé válik a strukturális gének transzkripciója. az E. coli lac régiója (szubsztrát - galaktóz, induktor - IPTG). Lásd a pozitív indukciót.
Negatív represszió - strukturális gének elnyomása, amelyben az effektor a szabályozó fehérjének azt a képességét adja, hogy az operátorhoz kapcsolódjon, megakadályozva a transzkripciót.
Kétértelmű átvitel. Lásd: Szuppresszió a riboszóma szintjén.
A nukleotidok helytelen párosítása - ritka nukleotidformák párosítása normál nukleotidokkal: A x - C, P - T, ahol A x és G x purinok ritka imino formában, és G - T x, A - C, ahol T x és Cx pirimidinek ritka enol formában.
Az inkompatibilitás azt jelenti, hogy az azonos inkompatibilitási csoportba tartozó rokon plazmidok nem képesek egyidejűleg létezni ugyanabban a baktériumsejtben.
A nonszensz kodonok olyan kodonok, amelyek nem felelnek meg egyik aminosavnak sem, és terminációs kodonként működnek a transzláció során (UAG - borostyánsárga, UAA - okker, UGA - opál).
reakciósebesség. Lásd Módosítások.
A nukleoid a baktériumok eukarióta magjának megfelelője, az RNS-t tartalmazó inkovírusok magja, amely RNS-ből és az azt körülvevő fehérjeköpenyből áll.
A nukleoproteinek nukleinsavak és fehérjék komplex komplexei.
A nukleoszóma az eukarióták kromoszómájának szerkezeti eleme, amely biztosítja annak stabilitását. Négy hisztonosztályból áll, amelyek gömb alakú testet alkotnak. A nukleoszóma magja két H4 hisztonmolekula tetramerje; kívül két molekula hiszton H2A és hiszton H2B (összesen 8 molekula). Nukleoszóma átmérője 10 nm. E struktúra köré egy legfeljebb 230 bázispár hosszúságú kettős szálú DNS-szegmens helyezkedik el, amely csaknem két fordulatot képez a nukleoszóma körül. A szomszédos nukleoszómák rövid DNS szakaszokkal kapcsolódnak egymáshoz.
Reverz transzkriptáz. Lásd Revertase.
A korlátozott transzkripció a genom tökéletlen leolvasásának esete, amikor olyan gének vannak megzavarva, amelyek termékei más gének aktivitásának megnyilvánulásához szükségesek. Például egy lizogén törzs indukciója során, amely λ-profág mutánst hordoz az N-génben, csak maga az N-gén íródik át a megfelelő mRNS-ekbe (iniciáció a P-promoteren).
Az in vitro kísérletekben, amikor az N génterméket adjuk a keverékhez, kiterjedtebb transzkripció figyelhető meg (a cIII, vörös, xis, int, cII, O gének leolvasása).
A nemhez kötött tulajdonságok olyan tulajdonságok, amelyek csak az egyik nemnél jelennek meg, vagy amelyek kifejeződése nemenként változik. Meghatározhatók mind az autoszomális, mind a nemi kromoszómákon található gének alapján. A juhoknál például a szarvasságot a H gén, a polledséget a h gén határozza meg. Ugyanakkor a kosoknál H > n, az anyajuhoknál pedig éppen ellenkezőleg, n > H. A H dominanciáját a hím nemi hormonok jelenléte határozza meg, ezért heterozigóta nőstényekben nem fordul elő.
Az egyértelmű gének nem allél gének, amelyek fenotípusosan ugyanúgy manifesztálódnak.
Az onkogének olyan fehérjéket kódoló gének, amelyek az eukarióta sejtek rosszindulatú átalakulását okozhatják.
Az ontogenezis az egyed egyedfejlődése, átalakulásainak összessége a születéstől (a tojás megtermékenyítése, a vegetatív szaporodási szerv önálló életének kezdete vagy az egysejtű anyai egyed osztódása) az élet végéig.
Az ontogenetikus adaptáció a szervezet azon tulajdonsága, hogy az egyedfejlődés során alkalmazkodni tudjon a változó környezeti feltételekhez. Feltételesen osztva szövetre (sejtes) és organizmusra.
Az ontogeketika (fenogenetika) a genetika egyik ága, amely az ontogenetika örökletes alapjait vizsgálja.
Az ontogenetikai módszer egy technikák összessége egy örökletes betegség hordozóinak tanulmányozására mind homozigóta, mind heterozigóta formában. Tartalmazza az anomáliák heterozigóta hordozóinak meghatározására szolgáló módszereket. Például a fenilketonuria gén heterozigóta hordozója meghatározható úgy, hogy fenilalanint fecskendezünk a vérbe, amelynek szintjét a vérplazmában határozzák meg.
Az operátor a DNS egy olyan szakasza, amelyet specifikus represszor fehérjék "felismernek", és szabályozzák az operonok vagy egyedi gének transzkripcióját. A lac rendszerben ez a represszor kapcsolódási helye és az mRNS szintézis kezdete. Az operon két állapotban lehet - nyitott és zárt. A kezelő nyitott, ha nincs represszor, és zárt, ha represszor van hozzá csatlakoztatva. Az operátor bezárása megakadályozza az adott operon összes szerkezeti génjének átírását. A λ bakteriofágnak két operátora van a cI géntől balra és jobbra, ezeket bal és jobb oldali operátoroknak nevezzük (OL , O R).
Az operon egy koordinált genetikai szabályozás rendszere, amely egy vagy több szerkezeti génből és a hozzájuk kapcsolódó megfelelő akceptor (szabályozó) génekből áll. Az E. coli-ban a lac operon a P promoterrel kezdődik, amely tartalmaz egy helyet, amelyhez a katabolit génaktivátor fehérje kapcsolódik, és egy helyet az RNS polimerázzal való kölcsönhatáshoz. A promotert az O operátor követi, amelyhez a represszor kötődik, majd a strukturális gének következnek. A lac operon terminátorral végződik, amely nonszensz kodonokat tartalmaz:
Az aminosavak sorrendjének meghatározása fehérjében. A 2-4-dinitrofluor-benzol (DNFB) reakcióját az amino- vagy N-terminálison található szabad α-aminosav-csoporttal alkalmazzák. A DNFB-vel való reakció az N-terminális aminosav (DNP) erős színű dinitrofenil-származékát eredményezi:
Ezt követően a polipeptid teljes savas hidrolízisen megy keresztül, és egyedi aminosavakra bomlik. A polipeptidet részleges hidrolízisnek vethetjük alá, ami különböző hosszúságú fragmentumokat eredményez. Mindkét esetben az N-terminális aminosav DNP-származékai könnyen izolálhatók kromatográfiával. Ezután ezek a fragmentumok teljes hidrolízisen mennek keresztül, és meghatározzák bennük az aminosav-összetételt. Több aminosav-maradékot tartalmazó átfedő oligomerek előállításához olyan enzimeket használnak, amelyek bizonyos helyeken megszakítják a polipeptidláncot. Például a tripszin megszakítja az arginin és a lizin kötéseit a következő aminosav-maradékkal a C-terminálisról:
kimotripszin - aromás aminosavak kötései; leucin-aminopeptidáz - az első peptidkötés az N-terminálison; karboxil-peptidáz - az első kötés a C-terminálisról. Mindkét utóbbi enzim felhasználásával, és a fehérjemolekula mindkét végén szekvenciálisan felbontva a kötéseket, a vizsgálat minden szakaszában meghatározható, hogy az aminosavak közül melyik szabad.
Nukleinsavak nukleotidszekvenciájának meghatározása. Ehhez szelektív többszörös hidrolízist, majd a kapott fragmensek elemzését alkalmazzuk. Hidrolízis a hasnyálmirigy-rivonukleáz által. - A hasnyálmirigy ribonukleáz az endonukleázokhoz tartozik, és az RNS molekulán belül azon a helyen bontja meg a foszfodiészter kötéseket, ahol a pirimidin nukleotid (C, U, T) a 3'-C oldalon található. Így megszakítja a b-kötéseket (X- a P b - X). A hidrolízis eredményeként nukleozid (és oligonukleozid) trifoszfátok szabadulnak fel:
Hidrolízis ribonukleáz TI-vel (takadiasztáz). Ez az endonukleáz megszakítja a b-kötéseket a foszfor és az 5'C között, ha a guanin a 3' pozícióban van:
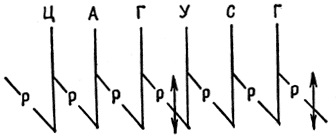
Hidrolízis kígyóméreg foszfodiészteráz által – ez az exonukleáz megtöri az RNS- és DNS-molekulákat a foszfátcsoport és az előző nukleotid 3S-e között. Egyenként szabadít fel 5'-nukleotidokat, a 3'-végtől kezdve (megszakítja az α-kötéseket):
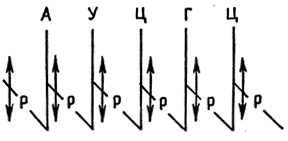
Hidrolízis szarvasmarha lép foszfodiészterázzal. Ez az exonukleáz nukleozid-3-monofoszfátokat szabadít fel RNS- és DNS-molekulákban, az 5'-végtől kezdve (megszakítja az α-kötéseket):
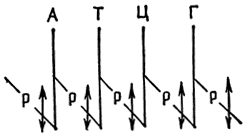
Origin (ori) - az a hely, ahol a DNS replikációja vagy átvitele megkezdődik.
Legyengített mutációk - E. sap mutánsok a relA gén károsodásával (angolul relaxed - gyengített RNS szintézis kontroll), ami aminosav éhezés során felszabadítja a sejteket az rRNS és tRNS szintézis szigorú kontrollja alól. Normális esetben a relA gén terméke, az úgynevezett szigorú kontrollfaktor, aminosav-éhezés során katalizálja a GDP-ből (ppG) és ATP-ből (pppA) egy szokatlan guanozin-3-difoszfát-5-difoszfát (ppGrr) nukleotid képződését. A normál törzsek „szigorúan” (szigorúan) szabályozzák az rRNS és a tRNS szintézisét. A gyengített kontrollú mutánsokból hiányzik a szigorú kontroll aktív faktora, ezért nem halmozódnak fel ppGrr-t az aminosav-éhezés során. A szigorú szabályozási faktor csak a 70S riboszómákkal együtt aktív, amelyek aminoacil-központjában a következő kodonnak megfelelő deacilezett tRNS-t hordoznak. Lehetséges, hogy a ppGrr felhalmozódása mindkét típusú RNS szintézisének leállítását okozza, megakadályozva például a polinukleotid láncok iniciációját azokban a promóterekben, ahol az rRNS és a tRNS transzkripciója normálisan beindul.
Kiválasztás - a különböző egyedek vagy egyedcsoportok utód elhagyásának eltérő valószínűsége. Az utódnemzés valószínűségét a szervezet számos tulajdonsága határozza meg: életképesség, a szaporodási kor elérésének sebessége, a szaporodási időszak időtartama, a keresztezési képesség és a termékenység. E tulajdonságok összességét az egyed környezeti feltételekhez való alkalmazkodóképességének nevezzük, és a genotípus határozza meg. A több összehasonlított csoport közül a magasabb alkalmasságát 1-nek, a többiekét az egy töredékének vesszük. Ha az aa egyedek utód elhagyásának valószínűsége 10%-kal kisebb, mint az AA és Aa egyedeké, akkor AA és Aa (w) alkalmassága 1, ebben az esetben w aa esetén 0,9. A szelekció intenzitásának kritériuma az összehasonlított csoportok alkalmasságának különbsége, amit S szelekciós együtthatónak nevezünk. Ebben a példában S = w AA - w Aa = 1 - 0,9 = 0,1.
Távoli hibridizáció - olyan hibridizáció, amelyben különböző fajokat és nemzetségeket keresztezik különböző fajok egyedi gének és kromoszómáinak kombinációival, és néha például alloploid hibridek és teljes genomok kombinációinak beszerzésekor, amely lehetővé teszi számukra, hogy egyesítsék az olyan formák tulajdonságait, amelyek szisztematikusan és biológiailag távol állnak egymástól. Meglehetősen nehéz távoli hibridizációt végrehajtani különböző okok miatt: a pollencsövek és a bibeszövetek összeférhetetlensége a növényekben, a nemi szervek szerkezetének és a szaporodási ciklusoknak való eltérése stb. előzetes oltások a szövetek vegetatív konvergenciája, pollenkeverékkel történő beporzás céljából. A távoli hibridek sterilitásának leküzdésére ígéretes módszer az amfidiploidok előállítása.
A nyitott leolvasási keret olyan nukleotidszekvencia, amely aminosavakat kódoló triplettek sorozatából áll, és nem tartalmaz transzlációs terminációs kodonokat. Egy ilyen szekvencia potenciálisan lefordítható fehérjévé.
A javítási hibák elsődleges mutációs léziók, amelyek a javítórendszerben és a kapcsolódó replikációs és rekombinációs rendszerekben előforduló enzimhibákból erednek. A hibák a spontán mutációk jelentős részét teszik ki.
Palindrom (palindrom szekvencia) - a DNS olyan szakasza, amelyben teljesen vagy csaknem azonos bázisszekvenciákat "olvasnak" mindkét irányban egy szimmetriaközpontból:
ABSSVA ABSSVA
Panmixia - különböző genotípusú heteroszexuális egyedek szabad keresztezése keresztezést megtermékenyítő szervezetek populációjában.
A partenogenezis az organizmusok ivaros szaporodásának egyik formája, amelyben a női csírasejtek megtermékenyítés nélkül fejlődnek.
Pachitén DNS - eukarióták DNS-e, amelynek szintézise a meiózis pachiténjében található, ami néhány már meglévő DNS-szakasz cseréjéhez vezet, és a reparatív szintézis típusának megfelelően halad. A pachytén DNS szintézise a kromoszómális DNS felett szétszórt régiókban megy végbe (gyakran többször is megismétlődik), azonos bázisszekvenciával. Ezek a szakaszok a genom 0,1 részét teszik ki. A pachytén DNS szintézisét hagyományos javító és replikációs enzimek végzik. Kivételt képez egy speciális endonukleáz (nikáz), amely csak a pachytenikus stádiumban jelenik meg.
Penetrance - egy adott gén alléljának megnyilvánulásának gyakorisága egy rokon organizmuscsoport különböző egyedeiben.
A peptidázok olyan proteolitikus enzimek, amelyek lehasítják a terminális aminosav-maradékokat a fehérje- és peptidmolekulákról.
A peptidil-transzferáz egy enzim, amely katalizálja a peptidkötés kialakulását, és az 50S riboszómában található. A peptidkötés kialakulása a formil-metionin-tRNS fmet és a riboszóma peptidil-, illetve aminoacil-centrumában található aminoacil-tRNS között megy végbe, és elősegíti a dipeptidil-tRNS képződését. Lásd: Közvetítés.
A peptidil-centrum az 50S riboszóma helye, amelybe a tRNS az aminoacil-centrumból költözik, amikor a következő aminoacil-tRNS megközelíti. Lásd: Közvetítés.
A peptidkötés az amidkötés egy fajtája, amely az egyik aminosav a-aminocsoportjának (-NH2) és egy másik aminosav α-karboxilcsoportjának (-COOH) való kölcsönhatásából jön létre.
Peptidek - szerves anyagok, amelyek maradékaiból azonosak vagy különféle aminosavak peptidkötés köti össze.
Elsődleges szűkület - a kromoszóma szűkítése, két karra osztása. A centroméra az elsődleges szűkület tartományában található. A centromer helyzetétől függően a kromoszómák metacentrikusak (egyenlő karok), szubmetacentrikusak (egyenlő karok) és akrocentrikusak (rúd alakúak).
A fehérje elsődleges szerkezete az aminosavak elrendezése egy polipeptidben. Lásd: Az aminosavszekvencia meghatározása fehérjében.
A génáramlás az egyedek populációból populációba való vándorlásával összefüggő gének (allélok) gyakoriságának változása.
A perliázok olyan hordozófehérjék, amelyek részt vesznek az anyagoknak a membránokon keresztül történő aktív szállításában.
Permutáció - a lókuszok általános rendjének ciklikus permutációi, például abvgdezikl orgzikl ...
A plazmagének a citoplazmában lokalizált örökletes faktorok, amelyek képesek az autoreprodukcióra és az örökletes információk továbbítására.
A plazmidok (episzómák) a baktériumok további körkörös kromoszómái, amelyek általában autonóm módon replikálódnak, és amelyek jelenléte nem szükséges a sejt túléléséhez. Egyes plazmidok beépülhetnek a bakteriális kromoszómába (F-faktor). A plazmidok hossza a bakteriális kromoszóma 0,05-1%-a. Az F-faktoron kívül léteznek R-faktorok, gyógyszerrezisztencia-faktorok (sztreptomicin, kloramfenikol, szulfonamidok), Col-faktorok, vagy kolicinogén faktorok (Shigella, Salmonella, Escherichia coli esetében), amelyek colicin termelődését okozó géneket tartalmaznak. , speciális fehérjeanyagok, amelyek képesek elpusztítani az azonos fajhoz tartozó baktériumokat, amelyek nem rendelkeznek ezzel a bakteriocinnal. Általában minden plazmid donor tulajdonságokat kölcsönöz a sejtnek.
Plazmon (plazmotípus) - a citoplazmában és sejtszervecskéiben lokalizált örökletes tényezők halmaza.
Plastidome - sejtplasztidok halmaza, mint örökletes információkat továbbító szerkezetek.
A pleiotrópia egy gén többszörös működése, egy gén azon képessége, hogy több tulajdonságra is hat.
Egy gén pleiotróp hatása egy gén különböző tulajdonságokra gyakorolt hatása. Ez azzal magyarázható, hogy egyetlen gén transzkripciós termékét több, egymással összefonódó növekedési és fejlődési folyamatban használják fel. Egy gén pleiotrópiáját a mutációi által okozott fenotípusos változások tanulmányozása tárja fel. Emberben a gén pleiotróp hatását számos génmutációra jellemző szindrómák (a fenotípus kóros elváltozásainak komplexumai) vizsgálatában találjuk. A domináns mutáció által okozott arachnodactiliában szenvedőknél az ujjak és lábujjak megnyúlnak, veleszületett szívhibák figyelhetők meg. Egy ritka örökletes betegség, a galaktoszémia demenciához, májcirrózishoz és vaksághoz vezet. Ez a tünetegyüttes a galaktóz-1-foszfát-uridiltranszferázt, a galaktóz (tejcukor) felszívódásához szükséges enzimek egyikét kódoló gén recesszív mutációjának következménye. A növényekben a gének pleiotróp hatását a klorofill szintézisét befolyásoló génmutációk példájával illusztrálhatjuk. Az ilyen mutációk amellett, hogy gyengítik a zöld színt, befolyásolják a növények növekedését, a levelek és virágok számát és méretét, valamint a vetőmag termőképességét.
A ploidia a többsejtű szervezet sejtjében vagy összes sejtjében található kromoszómakészletek száma.
Plusz-mínusz szál – az egyszálú fágok (FX174) DNS-t tartalmaznak, amelyet plusz-szálnak neveznek. Amikor a fág DNS (plusz szál) belép a gazdasejtbe, egy komplementer szál kezd ráépülni, amely templátként szolgál az új fág DNS képzéséhez, amelyet mínusz szálnak neveznek. Így a genetikai információ egy plusz láncba van zárva. Az RNS-tartalmú fágok (vírusok) típusuktól függően plusz láncot (poliovírus) vagy mínuszláncot (influenzavírus) tartalmaznak. Miután a plusz szál bejut a sejtbe, azonnal megkezdi az mRNS szerepét. A mínusz szálon először egy plusz szálat kell szintetizálni, mielőtt a benne található genetikai információt felhasználni lehetne.
A mutációk fokozása. Lásd: Lefelé irányuló mutációk.
A pozitív indukció a szabályozás olyan fajtája, amelyben a szabályozógén fehérjeterméke nem tiltja, hanem aktiválja a szintézist. Megfigyelhető az Escherichia coli katabolikus operonjában, amely az arabinóz asszimilációjára szolgáló enzimeket kódolja (igen - operon). A "szabályozó fehérje-arabinóz" komplex affinitást szerez az operon promóter részéhez, ahhoz kötődik, és aktiválja a szerkezeti gént.
A pozitív represszió egyfajta szabályozás, amelyben az operon munkáját aktiváló szabályozó fehérjét az effektor inaktiválja.
Nem - a szervezet jeleinek és tulajdonságainak halmaza, amely biztosítja részvételét az utódok szaporodásában és az örökletes információk átvitelében a következő generáció számára az ivarsejtek képződésén keresztül.
A polimerázok olyan enzimek, amelyek kis molekulatömegű anyagokból makromolekulák képződését katalizálják.
A szervek polimerizációja az a folyamat, amely a filogenezisben növeli az egyenértékű homológ képződmények számát a testben.
A polimeria egy kvantitatív tulajdonság több gén általi, egyértelmű hatású genetikai meghatározása. Az ilyen géneket polimernek nevezzük, és egyetlen betűvel jelöljük, amely a különböző gének számát jelzi (A 1 , A 2 , A 3 ). A polimerizmusnak két típusa van: kumulatív, amelyben egy tulajdonság megnyilvánulása a genotípusban lévő domináns gének számától függ, és felhalmozódásukkal növekszik, és nem kumulatív (például csirkékben a tollas lábak öröklődése, hüvely a pásztortáskában), amelyben az „egyedi hatású” gének határozzák meg a minőséget. Mindkét esetben bármelyik domináns gén jelenléte meghatározza domináns tulajdonság, és az aaaa forma recesszív fenotípusú. Mivel kumulatív polimerizmus esetén a tulajdonság a domináns gének számától függ, a felosztást a bizonyos számú domináns gént tartalmazó genotípusos osztályok gyakorisága is kiszámítja.
A polimer gének olyan nem allél gének, amelyek egy adott tulajdonságra ugyanolyan vagy közel azonos hatást fejtenek ki (gyakran kvantitatívak), amelyek additív hatást fejtenek ki. Lásd: Polymeria.
A polimorfizmus egy populációban számos genetikailag eltérő forma megléte, amelyek szaporodás közben szaporodnak. A heterozigóták populációban való jelenléte miatt megmarad a genetikailag és fenotípusosan eltérő egyedosztályok egy bizonyos aránya, amit kiegyensúlyozott polimorfizmusnak neveznek, például a társadalmi rovarok különböző formái közötti funkciók megosztása: méhek, hangyák, termeszek. . A polimorfizmus egy olyan mechanizmus, amely egy populációt egyetlen rendszerként tart fenn. A polimorfizmus biokémiai szinten is megnyilvánulhat. Lásd: Isoenzymes, Genetic Cargo.
A polinukleotid-foszforiláz egy enzim, amely ribonukleozid-difoszfátokat polimerizál. Nem igényel DNS-sablont.
A polipeptidek aminosavakból (6-10-től több tízig) épülő polimerek.
Polythecus - DNS-molekulák többszörös replikációja későbbi citotómia (sejtosztódás) nélkül, amelynek következtében óriási kromoszómák képződnek (például a rovarlárvák nyálmirigyeinek sejtjeiben).
Polifilia - egy adott szervezetcsoport eredete több olyan ősi csoportból, amelyek nem állnak szorosan rokonok között.
Nemi kromatin - a kromatin szakaszai, amelyek meghatározzák a különböző nemű egyének interfázisú magjainak különbségét, amelyek a nemi kromoszómák szerkezetének vagy működésének jellemzőihez kapcsolódnak. A nőstények magjainak 70%-ában, a hímek 5-6%-ában fordul elő. A nemi kromatinok jelenléte és száma az X kromoszómák számától függ. A nemi kromatinok száma eggyel kevesebb, mint a sejtben lévő X kromoszómák száma. Ez annak köszönhető, hogy egy kivételével minden X-kromoszóma spiralizálódik, és festődés után nemi kromatinként jelenik meg. A nemi kromatin meghatározását az orvosi gyakorlatban az egyén nemének megállapítása során használják, ami lehetővé teszi az interszexualitás nemének meghatározását a magzatban és az újszülöttben. A vizsgálathoz leukociták sejtjeit, az epidermisz bazális rétegét és a szájnyálkahártya kenetéből származó sejteket használják.
A nemi kromoszómák olyan kromoszómák, amelyekben a különböző nemű egyedek különböznek egymástól. A Drosophila melanogasterben 8 kromoszóma 4 párt alkot. Három kromoszómapár morfológiailag azonos, a negyedik pedig heteromorf. Ennek a párnak az egyik kromoszómája görbült és szubmetacentrikus (Y-kromoszóma), a másik (X-kromoszóma) akrocentrikus. A nőknek van egy sor nemi kromoszómája - XX, a hímeknek - XY. Lásd: Heterogametikus szex, Homogametikus szex.
A poláris mutációk nonszensz mutációk, amelyek csökkenthetik ugyanazon operon összes később leolvasott génjének szintetikus aktivitását. Az operátor és a mutáns gén között elhelyezkedő, azaz az előtte olvasott gének aktivitását nem érinti. A nonszensz mutáció poláris hatását az magyarázza, hogy normál esetben a riboszómák lezárják a teljes mRNS-molekulát, és kis területek szabadon maradnak az egyik gén terminációs kodonjai és a másik gén kezdeti kodonjai között. Abban az esetben idő előtti felmondás A transzkripció során nonszensz mutáció következtében az mRNS jelentősebb szakaszairól derül ki, hogy mentesek a riboszómáktól, jobban ki vannak téve a nukleázok hatásának, és természetesen a genetikai információ egy része elvész.
Lefelé irányuló mutációk - a lacP lókusz mutációi a BAC-cAMP komplex kötőhelyén, amelyek csökkentik a lac gének maximális transzkripciós sebességét (a lac operont tartalmazó DNS képessége csökken a jelzett komplexhez, ami csökkenéshez vezet a transzkripció beindulásának valószínűségében). A boost mutációk ellentétes hatást fejtenek ki.
Populációs módszer - olyan módszer, amely lehetővé teszi az egyes gének vagy kromoszóma-rendellenességek eloszlásának tanulmányozását az emberi populációkban. A géngyakoriság vizsgálata fontos az emberi örökletes betegségek elemzéséhez, az elszigetelt populációkban gyakori rokonházasságok következményeinek felméréséhez, valamint az emberi populáció genetikai történetének tisztázásához.
A populáció ugyanazon fajhoz tartozó egyedek összessége, amelyek közös génkészlettel rendelkeznek és egy bizonyos területet foglalnak el.
A fajtafajta az ember által mesterségesen létrehozott élőlények populációja, amelyek sajátos örökletes tulajdonságokkal rendelkeznek. A fajtán és fajtán belül minden egyednek hasonló öröklött rögzített tulajdonságai vannak: termőképesség, saját fiziológiai és morfológiai tulajdonságok komplexuma, azonos típusú reakció a környezeti tényezőkre. A fajta és fajta tulajdonságai a legjellemzőbb formában csak bizonyos természeti körülmények között jelennek meg.
Utócsökkentés. Lásd: Előredukció.
megfelelő operátor. Lásd a kezelőt.
A primer egy kis RNS-fragmens (50-200 nukleotid), amely egy újonnan szintetizált DNS-szakaszt indít el. Az RNS primereket egy „mobil promoter” indítja be. Az E. coliban ezt a funkciót a DNS B gén által kódolt B fehérje látja el, amely szekvenciálisan kötődik a késleltetett lánctemplát egy specifikus helyéhez, és kijelöli a primer inicializálásának helyét a primereket szintetizáló primáz (kezdeti RNS polimeráz) által. . Az E. coli primázt a dna G gén kódolja, a további meghosszabbítást E. coliban a DNS polimeráz III holoenzim (pol C és dna E, dna Z gének) végzi. A primert DNS-polimeráz I-gyel (pol A gén) távolítjuk el.
A citoplazma előre meghatározása - a citoplazma jellemzőivel kapcsolatos tulajdonságok öröklődésének esetei, amelyek az egyéni fejlődés folyamatában merülnek fel a környezeti tényezők hatására. Különbséget kell tenni az ontogenetikai és a genetikai előre meghatározottság között. Az ontogenetikai predetermináció a környezet által okozott változások a citoplazmában, amelyek nem stabilak és több generáció után eltűnnek. Például, ha a habrobracon nőstények petéit a megtermékenyítés előtt magas hőmérsékletnek teszik ki, az utódaik testszínének megváltozását eredményezi. Néhány generáció után normál hőmérsékleten ez a hatás eltűnik. Az ilyen változásokat, amelyek a kezdeti feltételekhez való visszatéréskor több generáció alatt elhalványulnak, hosszú távú módosításoknak nevezzük. A genotípus előre meghatározottságát az anyaszervezet genotípusa határozza meg. Példa erre a Limnea puhatestűben a héjcsavarodás irányának öröklődése, amikor az utódok fenotípusa az anya genotípusától függ, nem pedig a zigóták genotípusától, amelyekből kifejlődnek. Ugyanakkor a mendeli 3:1-es hasadás nem a második, hanem a harmadik generációban jelenik meg.
Preredukció - a nem-testvér kromatidák divergenciája az első meiotikus osztódásban. A második meiotikus osztódás eltérését posztredukciónak nevezik, és bármely gén és a centromer közötti keresztezés eredménye.
A shotgun elv (shot gun kísérletek) egy olyan módszer, amely az egyes gének klónozására szolgál egy szervezet genomjának DNS-ének fragmentálásával, és ezeknek a fragmentumoknak a baktériumsejtekbe való bejuttatásával bizonyos vektorok részeként. Ha elegendő számú baktériumklón található, amelyek a vizsgált DNS véletlenszerű fragmentumait hordozzák, akkor az egyik klónban megtalálható a kutató számára érdekes gén, amelyet azonosítani kell.
Fitness. Lásd a kiválasztást.
A proband az elsődleges beteg. Lásd a genealógiai módszert.
Progam ivarmeghatározás - a megtermékenyítés előtti ivarmeghatározás, amelyben a leendő egyed neme attól függ, hogy a nőstények kétféle tojást termelnek - nagy, gazdag citoplazmában és kicsi, citoplazmában szegény. A megtermékenyítés után az előbbiekből nőstények, az utóbbiakból hímek fejlődnek, például egyes férgekben, rotiferekben.
Prokarióták - egysejtű szervezetek amelyeknek nincs külön magjuk (főleg baktériumok).
Proximális gének - a kromoszómán vagy a bakteriális kromoszóma O-terminálisán kapcsolódó és szorosan elhelyezkedő gének.
Köztes öröklődés - a dominancia hiányának esetei, amikor egy hibrid egyedben egy tulajdonság mintegy közbenső helyet foglal el a szülők megfelelő tulajdonságai között.
A promóter az egyik akceptor gén, amelynek nukleotidpárjaiból álló szekvencia van, amelyet az RNS-polimeráz felismer, és amely hozzá kapcsolódik, majd az operon mentén mozog, átírva azt.
A protoplasztok olyan növényi sejtek, amelyek sejtfalát pektináz és celluláz pusztítja el. Szomatikus sejtek hibridizálására használják.
A prototrófok olyan baktériumok, amelyek képesek a szükséges összetett szerves anyagokat egyszerű anyagokból, például ásványi sókból szintetizálni, és minimális környezetben növekednek.
Feldolgozás - reakciók sorozata, amely a transzkripció és transzláció elsődleges termékeinek működő molekulákká történő átalakulásához vezet.
Ugró gének. Lásd transzpozonok.
A pszeudogének a globin szkriptonok (Ψβ1, Ψβ2, Ψα1) szegmensei, amelyek 80%-ban homológiát mutatnak bizonyos hemoglobinláncok cisztronjaival. Nem képeznek fehérjetermékeket, mivel sok olyan hibát hordoznak, amelyek megzavarják a transzlációs fázist.
Pszeudopoliploidia - a kromoszómák számának növekedése a genetikai anyag mennyiségének növekedése nélkül. Megkülönböztetik a fragmentációs pszeudopoliploidiát (a szomatikus sejtekben nagyszámú kis kromoszóma található, a csíravonal sejtekben pedig csak néhány nagy kromoszóma van, például a ló orsóféregében); agmatopseudopoliploidia (diffúz centromerek megjelenése, például spirogyra, egyes gombák és sok rovar esetében); kis kromoszómák fúziójából származó pszeudopoliploidia. Az utóbbi típusú pszeudopoliploidia nagy kromoszómák megjelenéséhez vezet különböző számban a különböző egyedekben, amelyek poliploid sorozatnak tekinthetők, például a saprogyra esetében. Egyes növényekben az úgynevezett B kromoszómák megjelenését figyelték meg, amelyek általában az A kromoszómák töredezettségének az eredménye. A B-kromoszómák szerepe nem tisztázott.
A biokémiai reakciók elágazása- különböző végtermékek előállítása független módon egy prekurzorból:
A fehérjék feloldása. Lásd: Helicases.
Hasadás - eltérő genotípusú egyedek (sejtek) hibridjének megjelenése az utódokban vagy az utódok genotipikusan meghatározott különbsége egy tulajdonság megnyilvánulása szempontjából.
Visszaállítás. Lásd mutáció.
A revertáz (reverz transzkriptáz) az onkogén RNS-tartalmú vírusok enzime, amely az úgynevezett reverz transzkripciót, azaz a provírus DNS szintézisét hajtja végre egy vírus RNS templáton. A szintézis során RNS-DNS hibrid képződik, majd a DNS-lánc a DNS-függő DNS polimeráz hatására replikálódik, és a létrejövő kettős DNS-lánc további replikáción megy keresztül. A reversetas által szintetizált vírus DNS beépül a fertőzött sejt genomjába.
Szabályozó - egy represszor szerkezetét kódoló gén, amelynek feladata az operon transzkripciójának szabályozása.
Ritka nukleotidok (kisebb) – szokatlan bázisú nukleotidok, például inozin (I), 1-metil-guanilsav (Gm), 1-metil-inozin (Im) és dimetil-guanilsav. A metilcsoportok jelenléte megakadályozza a komplementer párok kialakulását. Ritka bázisok közé tartozik a pszeudouridil (Ψ) is, amelyben az uracil pirimidingyűrűje nem az 1-es pozícióban lévő kötésen, hanem az 5-ös pozícióban lévő szénen keresztül kapcsolódik a ribózhoz, a ribotimidilsav (T), amelynek az uracilhoz hasonló bázisa van. és egy metilcsoportot tartalmaz az 5. pozícióban.
Újrakezdés - a prokarióta sejtek azon képessége, hogy újraindítsák a fehérjeszintézist, amit az ebből eredő nonszensz mutáció megszakít. Nemcsak az AUG (Met) kodon, hanem más kodonok is szolgálhatnak újrakezdési jelként. Az eukariótákban nem találtak reiniciációt.
Tiltott (vagy szabálytalan) rekombináció – nem homológ cseréket (transzlokációkat, inverziókat és egyenlőtlen keresztezési eseteket) magában foglaló rekombináció, mint például a fág és a bakteriális DNS közötti rekombináció homológia hiányában. Az ilyen illegitim rekombináció a λ-profág integrációját és kizárását eredményezheti, ami hibás transzdukáló fágokat eredményez, különböző mennyiségű fág DNS-sel.
Helyspecifikus rekombináció – rekombináció okozta. integrációs fehérje, amelyet a λ fág int génje kódol. Ez a fehérje specifikusan kötődik egy speciális integrációs helyhez, amely a β fág genom b2 régiójában található, és csak ezen a helyen idéz elő keresztezést a bakteriális és a fág kromoszómák között.
Replikáz – RNS-függő RNS polimeráz.
A replikon a genom egy olyan régiójának replikációs folyamatának egysége, amely egyetlen iniciációs pont irányítása alatt áll.
A repliszóma fehérjék komplexe, amely a replikációs villában képződik, és a DNS-replikáció minden szakaszában a normál áthaladást végzi. A replizóma komponens lehet DNS-polimeráz I, II, III, DNS-függő RNS-polimeráz, Okazaki-fragmensek képződésében részt vevő RNS, ezeket a fragmentumokat összekötő polinukleotid-ligáz; topoizomeráznak nevezett enzimek, amelyek funkciója a DNS szuperspirálozás mértékének megváltoztatása; olyan enzimek, amelyek megolvasztják a DNS-t, azaz elválasztják a komplementer szálakat, amelyek biztosítják a DNS feltekercselését; láncainak szétválasztása; oltó RNS-fragmens (primer) szintézise, majd Okazaki-fragmensek képzése; RNS primer eltávolítása; a kialakult egyhélix hézag kitöltése és az Okazaki-töredékek kovalens kapcsolata.
Represszió - a génaktivitás elnyomása, leggyakrabban a transzkripció blokkolásával.
A represszor olyan fehérje, amely elnyomja egy vagy több olyan gén transzkripcióját, amelyek szorosan kapcsolódnak egy operonhoz vagy egy kromoszómán szétszóródnak.
A restrikció azt jelenti, hogy egy adott fágra általában érzékeny baktériumtörzsek nem képesek támogatni annak növekedését. A korlátozott fág ezért korlátozott számú gazdaszervezettel rendelkezik. Lásd: Gazdamódosítások.
A retrovírusok az RNS-vírusok családját alkotják (TMV, HT LVI és 2, LAV/HTLV3).
Retroinhibíció - ennek a metabolikus láncnak az első enzimének gátlása a végtermék által, amely általában kis molekulatömegű (aminosav). A biokémiai utak elágazása és az izoenzimek hiánya esetén az ún. koordinált retroinhibíció figyelhető meg. Például a Baccilus nemzetségbe tartozó E. coli-val ellentétben három aszpartát-kináz helyett csak egy vesz részt a lizin, treonin és izoleucin szintézisében, és retrogátlása csak mindhárom aminosav egyidejű feleslegével történik. Lásd az izoenzimeket.
Recesszívség - az egyik allél fenotípusos megnyilvánulásának hiánya heterozigóta egyénben.
Kölcsönös keresztezések - olyan keresztezések, amelyekben élőlények domináns és recesszív vonások anyaként és apaiként egyaránt használják.
Hasnyálmirigy ribonukleáz. Lásd: Nukleinsav-szekvenálás.
A riboszóma egy citoplazmatikus szerkezet, amelyen polipeptidek szintetizálódnak.
A DNS-függő RNS-polimeráz egy olyan enzim, amely katalizálja a transzkripciót (RNS-szintézist). Templátként egyszálú vagy denaturált DNS-t használ, amelyen RNS-t szintetizál. Ebben az esetben a vetés nem szükséges. Az enzimnek mind a négy 5'-trifoszfát-tribonukleozidra szüksége van, és ötféle alegységet tartalmaz: α, β, β", σ, ω. Az α2ββ"ω szerkezetet "core-enzimnek" vagy "minimális enzimnek" nevezik. Az αββ"ωσ komplex holoenzim. A magenzim katalizálja a nem specifikus RNS szintézist, amely a DNS-templát bárhol megindulhat. Egy σ faktor hozzáadása szelektív transzkripciót eredményez. Csak a promóter régióban kezdődik. Lásd: RNS szintézis.
RNS polimeráz RNS-függő - replikáz.
Hely - egy pontmutáció helye a gén rekombinációs térképén; a fágtérképeken néha a kromoszóma egy teljes régiójának megjelölésére használják, például a λ fág b2-ét, az int helyet, azaz az integrációs helyet. . Ha két allélmutáció rekombinálódik egymással, akkor különböző helyeken lokalizálódnak.
A szatellit DNS az eukarióta DNS része, amely több nukleotidból álló, 150-300-szor ismétlődő rövid szekvenciából álló klaszterekből áll. Az eukarióta DNS jelentős részét a szatellit DNS teszi ki: Drosophila 4-12%-át, emberben akár 15%-át, főként centromer heterokromatin és telomer régiókban lokalizálódik.
A kiegyensúlyozott letálisok olyan gének, amelyek recesszív halálos hatást fejtenek ki. Jelenlétük a populáció genotípusában megmarad természetes kiválasztódás, mert a gének működésének pleiotrópiája miatt ezek az allélok heterozigóta állapotban bizonyos körülmények között bizonyos pozitív hatást válthatnak ki. Azokat a vonalakat, amelyekben a recesszív letálisok heterozigótasága automatikusan megmarad, kiegyensúlyozott letális rendszereknek nevezzük.
Kiegyensúlyozott polimorfizmus. Lásd: Genetikai homeosztázis.
Svedberg egység (S) - ülepedési egység, a képlet határozza meg
ahol ω 2 - szöggyorsulás; dx/dt - részecskeelmozdulás időegységenként; ω 2 x annak a gravitációs mezőnek a jellemzője, amelyben a részecske mozog az ultracentrifugálás során. Néha helyettesíthető g - szabadesés gyorsítással.
A túlzott dominancia egy tulajdonság erősebb megnyilvánulása egy zigótában, mint egy adott allélpár bármelyik homozigótájában (AA és aa).
Szekvenálás (az angol szekvencia - szekvencia) - az RNS vagy DNS nukleotidszekvenciájának meghatározása.
A szexdukció a gének egyik baktériumsejtből a másikba történő átvitelének folyamata egy autonóm nemi faktor (F" = lac) segítségével.
A testvérek ugyanazon szülők leszármazottai.
Szingamikus nemi meghatározás. - A leendő egyed nemét a zigóta genotípusa határozza meg, és nem függ a környezeti feltételektől. A legtöbb kétlaki élőlényben előfordul.
A síró macska szindróma egy örökletes betegség, amelyet az ötödik emberi kromoszóma rövid karjának törlése határoz meg.
A szinkaryon két haploid mikronukleusz fúziójának terméke, amelyek csillósokban meiózison mentek keresztül.
A genetikai markerek megmentése - az egyik szülő genetikai markereinek megnyilvánulása rekombináns utódokban, ha elnyomták őket a szülői genomban.
A távtartók röviden elválasztó nukleotidszekvenciák a promoter és a strukturális gén között. Az E. coli 37 bázispárral rendelkezik. Nem átíródó régiók formájában ismétlődő ismétlődésük esetén rRNS gének között fordulnak elő. Az eukarióta génekben a távtartók olyan szakaszok, amelyek elválasztják a gén átírt részeit - szkriptonokat.
Specifikus kombinálási képesség - az önbeporzó vonal megnövekedett értéke egy adott keresztkombinációban. Sok vonal egymással való keresztezésével határozható meg.
Szomatikus sejtek - többsejtű szervezetek szövetsejtjei, amelyek nem szexuálisak.
Illesztés. Lásd Intron.
A sporofita a növények életciklusának ivartalan diploid nemzedéke. Megtermékenyített tojással kezdődik, és spórák képződésével végződik.
Fajta. Lásd Fajta.
Az RNS-szintézis szigorú ellenőrzése - az rRNS és a tRNS szintézisének befejezése aminosav-éhezés során. A rel A gén (E. coli) szabályozza. Lásd: Gyengített mutánsok.
A felülfertőzés a korábban új fágrészecskékkel fertőzött baktériumsejt további fertőzése.
Szuppresszió riboszóma szinten – a sztreptomicin jelenléte egyes kodonok félreolvasását okozza, ami például a leucin, tirozin, szerin felvételéhez vezet a fenilalanil-kodonra válaszul.
A szupresszor mutáció olyan mutáció, amely az elsődleges mutáció következtében elveszett funkció teljes vagy részleges helyreállítását okozza.
Szupresszor tRNS - tRNS, amelynek antikodonja egy mutáció eredményeként képes volt felismerni az mRNS egyik vagy másik nonszensz kodonját és szignifikánsnak olvasni. Néha az antikodon változatlan marad a szupresszor tRNS-ben. Például a szupresszor tRNS trp-ben az antikodon (ACC) nem változik, de a 24. pozícióban, az antikodontól 1 nm-re nukleotid változás történik. Nyilvánvalóan ennek a pótlásnak köszönhetően a tRNS kvaterner szerkezete annyira megváltozott, hogy antikodonját nemcsak az UGG kodon (triptofán), hanem a termináló UGA kodon (opál) is felismeri.
Olvasási eltolás-szuppresszorok - az akridin festék által kiváltott frameshift mutációk elnyomása (deléciók, inszerciók). Léteznek intra- és intergenikus szuppresszorok. Az intragenikus szuppresszorok hatása abban áll, hogy az eredeti frameshift mutáció helyétől (deléció, inszerció) nem messze inszerció, deléció (ellentétes előjelű mutáció) következik be, ami a teljes cisztronban a normál leolvasási keret visszaállításához vezet. , kivéve a mutációk közötti területet. Az intergénikus szuppresszorok olyan szuppresszor mutációk, amelyek az eredeti mutációtól távol eső lókuszban lokalizálódnak. Ezen szuppresszorok egy része a baktérium tRNS-éből származik. Egy ilyen szupresszor tRNS visszaállítja a leolvasási keretet, ha a szintetizált polipeptid átjutott egy kereteltolásos mutáció helyén (például egy beépített bázison), és a mutáns tRNS nem 3, hanem 4 nukleotiddal halad előre.
A gének kapcsolódása egy olyan jelenség, amely a gének csoportonkénti együttes átviteléből áll, ha ugyanazon a kromoszómán helyezkednek el. A csatolás lehet teljes vagy hiányos. A kapcsolódás mértéke az átkelés valószínűsége.
A nemi kötődés olyan tulajdonságok öröklődése, amelyek génjei a nemi kromoszómákon lokalizálódnak. A nemhez kötött vonásokat a kölcsönös keresztezések során mindkét nem eltérő szegregációja tárja fel. Az 1:1 arányú ivarosztódást az ivarkromoszómakészlet szerint kétféle ivarsejt képződése magyarázza egy heterogametikus ivar által az ivarsejtek egyik típusában egy homogametikus ivar által.
A telomerek a kromoszóma végei.
Lefagyott esetelmélet - ezen elmélet szerint a genetikai kód szerkezete véletlenszerű események hatására alakult ki, de miután az eredeti sejtben, amely minden élő szervezet közös őse, megállapították a kodonok jelentését, további evolúciós eltérések következtek be. A kód lehetetlenné vált, mivel minden olyan mutáció, amely megváltoztatja a kodon és az aminosav megállapított megfelelését, halálos lehet.
Terminátor (terminátor kodon) - egy DNS szegmens, amely stop jelként szolgál, amely megállítja az RNS polimeráz fejlődését, az operon átírását. Általában több ismétlődő nonszensz kodonból áll.
Szintrofizmus-teszt – egyes mutánsok azon képességének vizsgálata, hogy kompenzálják az anyagcsere-hibákat és stimulálják más mutánsok szaporodását egy közös tápközegben a felhalmozódó metabolitok feleslegének diffúziójával. Felhasználható az anyagcsere utak lépéseinek sorrendjének megállapítására.
A tetrad-analízis egy olyan módszer, amely lehetővé teszi annak bizonyítását, hogy a Mendel-hasadást a meiózis mechanizmusa határozza meg, és ez nem statisztikai, hanem biológiai mintázat. A sütőélesztőnek vörös és fehér telepei vannak. Ezeket az alternatív karaktereket egy allélpár határozza meg: A - fehér és - a kolónia piros színe. Az ivarsejtek fúziója egy diploid Aa zigótát eredményez. Hamarosan meiózisba lép, amelynek eredményeként haploid spórák tetrádja képződik az aszkuszban. A hasadás meghatározásához az aszkusz minden spóráját külön-külön táptalajra vetjük. A kialakult négy kolónia közül kettő fehér és kettő vörös, azaz 1A:1a hasadás figyelhető meg. Hasonló eredményt kapunk bármely két másik jellemző vizsgálatával. Ez a hasadás a meiotikus osztódás eredménye.
A traizáció egy olyan mutáció, melynek eredményeként a purinbázist egy purinbázis, a pirimidinbázist egy pirimidinbázis (A-T → G-C) helyettesíti.
Traissersion - egy mutáció, amelynek eredményeként a purinbázist pirimidin, a pirimidint pedig purin helyettesíti (A-T → T-A).
A transzgekózis a génsebészet egyik módszere. Ez egy genomból izolált gének vagy mesterségesen szintetizált gének kísérleti átvitele egy másik genomba. Három egymást követő műveletből áll: egy gén izolálása vagy szintézise, vektorba foglalása és a benne lévő gént tartalmazó vektor sejtbe juttatása.
A transzgresszió olyan gének összesítő hatása, amely egy tulajdonság növekedését vagy csökkenését okozza.
Transzdetermináció - egy szerv kezdetleges fejlődési irányának hirtelen változása hormonális rendellenességek miatt. A transzdetermináció oka lehet mind a homeotikus mutációk, mind a korongprimordiumok átültetése rovarokban.
A transzdukció egy DNS-fragmens átvitele egy donor sejtből a recipiens sejtbe vírus (mérsékelt égövi fág) segítségével. Tegyen különbséget az általános és a specifikus transzdukció között. Az általános (nem specifikus) transzdukció olyan transzdukció, amelyben a baktériumokban szaporodó fág képes befogni a bakteriális DNS bármely szakaszát, és átvinni egy másik, erre a fágra fogékony baktériumba. Általában 1-3 gén kerül átadásra. Egy transzdukáló fágrészecskében a genomjának egy részét a donor bakteriális DNS fragmentuma helyettesíti, és az ilyen részecske nem szaporodik, nem pusztítja el a sejtet, és nem teszi lizogénné. Specifikus (korlátozott) transzdukció - transzdukció, amelyben a donor kromoszóma egy bizonyos régiója átkerül a profág genom egy bizonyos lókuszhoz való kapcsolódása miatt. Például a λ fág lac operon melletti kapcsolódási helye és a hasítás során keletkezett úgynevezett defektív Idg fágok, amelyek genomjuk ~30%-ával rendelkeznek, hordozzák a gal géneket (az f80 fág a try géneket hordozza). Előfordul, hogy a transzdukáló fág genomja nem integrálódik a gazdakromoszómába, a sejt citoplazmájában marad, és osztódása során a két leánysejt közül csak az egyikbe jut be. Ezt a jelenséget fordított transzdukciónak nevezik.
A transzkripció a genetikai információ DNS-molekulából RNS-molekulává történő átírásának folyamata, vagyis az mRNS szintézise.
A transzlokáció egy gén vagy egy kromoszóma szegmens mozgása a genom egyik lókuszáról a másikra.
A transzláció a genetikai információ fordítása az mRNS-ben lévő nukleinbázisok nyelvéről a fehérjében lévő aminosavak nyelvére, azaz fehérjeszintézis.
A transzpozíció a genetikai anyag egy töredékének mozgása a sejtgenom nem homológ régiói között.
A transzpozonok (transzponáló elemek) kisméretű DNS-fragmensek, amelyek a kromoszómába integrálódva, annak mentén mozogva tartalmaznak a transzpozíciós rendszeren kívül egyéb funkcionális célú géneket (antibiotikumrezisztencia, anyagcsere-folyamatokat irányító gének).
A transzfekció a baktériumsejtek fertőzése, amikor azokat DNS-tartalmú vírusokból izolált tisztított DNS-készítményekkel kezelik, melynek eredményeként a sejtben új generációs virionok képződnek, mintha egy teljes értékű vírussal fertőződtek volna meg.
Három faktoros kereszt - genetikai keresztezések három különböző genetikai markert használva (például + /a + /b + /c × a/a in / in c / c).
A triszómia egy kariotípus-változás, amelyben egy diploid halmazban egy vagy több kromoszóma három kópiában jelenik meg.
A transzformáció a gének sejtek közötti átvitele DNS-donor segítségével. A befogadó sejtek csak az életciklus egy bizonyos szakaszában alakulnak át, amikor fogékonyak a transzformáló DNS-re, az úgynevezett kompetens sejtek. A donor DNS egy fragmentuma, amely behatolt a recipiens sejtbe, kettős keresztezéssel bekerül a kromoszómájába, kicserélve a megfelelő géneket. Megkülönböztetni az intraspecifikus és interspecifikus transzformációt.
Mérsékelt öv fág - olyan fág, amely képes a baktériumsejt lizogenizálására, profág állapotba kerülve, behatolni a gazda DNS-ébe (λ fág) vagy a citoplazmában maradni (PI fág).
Feltételesen letális mutációk - bizonyos körülmények között halálos mutációk: auxotróf - a szükséges növekedési faktorok hiányában; hőmérséklet érzékeny - emelt hőmérsékleten.
Helper fág - normál fágrészecskék (genomok), amelyek hozzájárulnak a hibás transzdukáló fágok (genomok) kialakulásához.
A szigorú szabályozási faktor az E. coli rel A génje által szabályozott enzim. Szabályozza az rRNS és a tRNS szintézisét, leállítja azt aminosav-éhezéssel. Lásd az attenuált mutációkat.
Elongációs faktorok - EF - Tu, EF - Ts és EF - G elongációs faktorok, amelyek szükségesek az aminosavak polipeptidláncokká történő összeállításához. Nem a riboszómák szerkezeti alkotóelemei, és csak a fehérje-összeállítás egy bizonyos szakaszában kötődnek hozzájuk.
Fenogenetika. Lásd: Ontogenetika.
Fenotípus - egy szervezet összes jelének és tulajdonságának összessége, amely genetikai szerkezetének (genotípusának) és a külső környezet kölcsönhatásának folyamatában alakul ki. A fenotípusban soha nem valósul meg minden genetikai lehetőség, vagyis az egyes egyedek fenotípusa csak egy speciális esete genotípusa bizonyos fejlődési feltételek melletti megnyilvánulásának.
A fenotípusos gyök az organizmus genotípusának az a része, amely meghatározza a fenotípusát. Az AABB, AaBB, AaBb, AABb, AaBb genotípusoknál a fenotípusos gyök A-B- lesz.
A fluktuációs teszt egy módszer a baktériumsejtekben bekövetkezett változások genetikai (mutáns) természetének bizonyítására, összehasonlítva e tulajdonságok előfordulási gyakoriságát (fluktuációját) független tenyészetekben (a különbségeknek statisztikailag szignifikánsnak kell lenniük).
Okazaki-fragmensek - a replikáció során az újonnan szintetizált DNS egy úgynevezett késleltetett szálon nagyszámú kis fragmentumból áll, amelyek körülbelül 1000 (zukariotákban körülbelül 200) nukleotidot tartalmaznak.
Freemartinok - emlősökben a heteroszexuális ikrek fejlődése során néha az egyikük neme megváltozik az embriogenezisben. Tehát a szarvasmarha heteroszexuális ikreinél a bikák normálisan fejlődnek, és az üszőkről kiderül, hogy interszexuálisak. Az ilyen állatokat freemartinoknak nevezik. Általában terméketlenek. Az ilyen változásokat az okozza, hogy a herék korábban kezdik el kiválasztani a férfi hormonokat a véráramba, mint a petefészkek.
Chiasma – X-alakú szerkezet, amely a homológ kromoszómák divergenciájának kezdete utáni átkelés eredményeként jön létre a meiózis I. profázisában.
A kromatin DNS fehérjékkel alkotott komplexe, amely egy kromoszóma, amely az interfázisú magban dekompaktált.
A kromoszómák a sejtmag nukleoprotein fonalas szerkezetei, amelyek affinitást mutatnak a fő festékhez. A mitózis és a meiózis során megfigyelték és azonosították. A kromoszómák fő axiális komponense egy óriási folyamatos DNS-molekula, amely lineáris sorrendben tartalmazza a géneket és a genetikai szabályozó szekvenciákat.
A cisz-transz teszt egy genetikai elemzési módszer, amely lehetővé teszi annak meghatározását, hogy két recesszív mutáció, amelyek fenotípusos megnyilvánulása hasonló, azonos vagy különböző génekhez tartozik-e. A cisz-transz teszt a gén mint funkcióegység elgondolásán alapul. Egy kettős heterozigóta a+/+b típusban ugyanabban a génben két mutáció mutagén fenotípust eredményez, ha transz konfigurációban van, és vad típust, ha cisz konfigurációban vannak.
A cisztron a DNS funkciójának egysége, amelyet a cisz-transz teszt határoz meg. A kifejezést a gén szinonimájaként használják annak a DNS-szekvenciának a meghatározására, amely egy polipeptidet koordinál.
A törzs egy (egy sejtből származó) poloska, eredetét tekintve mikroorganizmusok kultúrája, melynek genetikai specifitását szelekció tartja fenn.
Exonok - kódoló szekvenciák az eukarióták szerkezeti génjeiben; jelen van az érett DNS-ben.
Expresszivitás - egy genetráns gén expressziójának mértéke.
A génexpresszió a DNS-ben kódolt genetikai információ megvalósítása az mRNS transzkripciója és transzlációja révén.
Endogén - a bakteriális kromoszóma egy része, homológ a genom egy fragmentumával (exogén), amely a donortól a recipienshez kerül a képződés során.
Az episzóma egy plazmid, amely képes baktériumokat integrálni a kromoszómális DNS-be.
Episztázis - egy gén expressziójának elnyomása egy másik, nem allél gén által.
Az eukarióták olyan organizmusok, amelyek sejtjei határozottan osztódnak magra és citoplazmára. Az eukarióták lehetnek egysejtűek vagy többsejtűek.
A pozícióhatás egy gén megnyilvánulásában bekövetkező változás, amely a kromoszóma-átrendeződések során szokatlan helyre tolódik.
n1.doc
Genetikai szótár
ABERRÁCIÓ KROMOSZOMÁLIS - a kromoszóma szerkezetének átrendeződése bármilyen változással összefüggő. A kromoszómamutációk bármelyik típusának általánosított neve: osztódások, transzlokációk, inverziók, duplikációk. A kifejezés néha genomi mutációkat is jelent (aneuplodia, triszómia stb.).
ABRAHIA - felső végtagok hiánya.
AGENESIA (aplasia) - egy szerv vagy annak egy részének teljes veleszületett hiánya.
AGIRIA (lissencephaly) - barázdák és csavarodások hiánya az agyféltekékben.
AGLOSSIA - nyelvhiány.
AGNATIA - az alsó állkapocs apláziája
A GÉNEK MŰKÖDÉSÉNEK ADADITIVITÁSA - a nem allél gének kölcsönhatásának egy típusa, amikor a gének egy csoportjának általános fenotípusos hatása egy tulajdonságra megegyezik mindegyikük hatásának összegével.
AKROCENTRIKUS KROMOSZÓMA – az egyik, amelyben a centroméra az egyik végének közelében helyezkedik el, míg a kromoszóma egyik karja hosszú, a másik rövid.
Acrocephaly - magas koponya.
ALLÉL - egy gén formái, amelyek fenotípusos különbségeket okoznak, és homológ kromoszómák homológ régióiban lokalizálódnak. A keresztezés során az allélok a monohibrid hasítás szabályai szerint mendeliak, és direkt és fordított mutációkkal egymásba fordulhatnak. Az ivarosan szaporodó szervezetek allélizmusának bizonyítékát keresztezéssel végzett tesztelés útján nyerik. Sok gén (lókusz) esetében csak két allél ismeretes, amelyek közül az egyik, a „vad típusú allél”, a második allélt többnyire uralja. Azonban gyakran találkozunk több allél sorozatával, pl. allél gének sorozata, amelyek 3-20 vagy több különböző fenotípust okoznak. Az allélkapcsolatok, különösen a dominanciaviszonyokhoz kapcsolódóak, megváltoztathatók a genotípusos környezet megváltoztatásával. Egy gén fenotípusos megnyilvánulása elsődleges hatásának és más génekkel való kölcsönhatásnak az eredménye. A hasonló enzimek képződését vagy hasonló szintézisfolyamatokat szabályozó allélek eltérőek lehetnek a megnyilvánulásukhoz szükséges optimális hőmérséklet, pH vagy szubsztrát-szubsztrát feltételek tekintetében. Ha egy allélpárra heterozigótaság mellett csak az egyik jelenik meg, pl. a hibrid csak az egyik homozigóta szülővel mutat fenotípusos hasonlóságot, akkor a manifeszt allél ún. uralkodóés a másik - recesszív.
ALLÉL - egy gén két vagy több alternatív formájának egyike, amelyek mindegyikét egyedi nukleotidszekvencia jellemez.
Az allél gének lehetnek domináns, recesszívÉs kodomináns. A génpenetráció mindkét nemnél egyformán teljes vagy hiányos lehet, és változhat vagy korlátozódhat az egyik nemre.
ALOPECIA- tartós vagy átmeneti, teljes vagy részleges hajhullás.
ALFA-FETOPROTEIN (AFP)- újszülött, terhes nő magzatának vérében, valamint a magzatvízben található embrionális fehérje.
AMÉLIA- a végtagok teljes hiánya.
AMINOSAVAK- fehérje monomerek.
AMNIOTIC SZÁLAK(Simonar szálak) - az amnion malformációja szövetszálak formájában, amelyek a méhen belül haladnak át, és összekötik a placenta gyümölcsfelületének különböző részeit a magzat felületével.
AMNIOCENTÉZIS- genetikai anomáliák prenatális diagnózisának módszere. A magzatvíz zsák átszúrásával állítják elő, hogy magzati sejteket tartalmazó magzatvizet nyerjenek.
ERŐSÍTÉS- egy további gén képződése.
ELEMZÉSI KERESZT- heterozigóta keresztezése recesszív homozigótával (elemző egyeddel), ami lehetővé teszi a hibridben képződött ivarsejtfajták számának meghatározását.
ANAFÁZIS- a mitózis vagy meiózis egyik szakasza, amelynek során a kromoszómák a sejt ellentétes pólusaira térnek el.
ANEUPLOIDIA- olyan jelenség, amelyben a sejtek kiegyensúlyozatlan kromoszómákkal rendelkeznek. Ebben az esetben a fajspecifikus készletből egy vagy több kromoszóma hiányzik, vagy további másolatok képviselik őket, pl. a kromoszómák csökkent vagy megnövekedett száma nem a haploid (re) többszöröse. Az aneuploid formák közé tartozik a monoszómia (2p- 1), triszómia (2n + 1 + 1) és egyéb aneuploid formák, amelyek az egyes kromoszómák anafázisban történő elvesztése, a kromoszómák szétválasztásának hiánya, a többpólusú mitózisok miatt léphetnek fel a kromoszómák helytelen eloszlásával a leánysejtek között.
ANIRIDIA- az írisz hiánya.
ANKYLOBLEPHARON- a szemhéjak széleinek összeolvadása nyálkahártyával borított összenövésekkel.
ANOMÁLÁD- másodlagos többszörös anomáliák, amelyek egy ismert vagy feltételezett anomáliából vagy egy mechanikai tényező hatásából erednek.
RENDELLENESSÉG- rendellenesség, szabálytalanság, eltérés a normától.
ANORCHIZMUS- here agenesis.
ANOTIA- a fülkagyló apláziája.
ANOPHTALMIA- Egy vagy mindkét szemgolyó hiánya.
ANTIGÉN- idegen fehérje molekula, amely egy antitest szintézisét indukálja.
ANTIMONGOLOID SZEMSZEKCIÓ- a palpebralis repedések leengedett külső sarkai.
VÁRAKOZÁS- Az egyre koraibb megnyilvánulásra való hajlam a következő generációkban.
ANENCEFALY- Az agyféltekék hiánya (fejlődési rendellenesség).
APPLASIA- lásd agenesis.
APODIA- lásd Acheiria.
ARACHNODACTYLY- szokatlanul hosszú és vékony ujjak ("pók").
ARINENCEPHALIA- a szaglóhagymák, barázdák, traktusok és lemezek apláziája.
ARTROGRIPOZIS- az ízületek veleszületett kontraktúrái.
ASZORTÍV HÁZASSÁGOK- azok, amelyekben a házastárs egy vagy több alapon történő kiválasztása nem véletlen.
ATRESIA- csatorna vagy természetes nyílás teljes hiánya.
AUTOSZÓMA domináns ÖRÖKSÉG- az öröklődés olyan típusa, amelyben egy, az autoszómában lokalizált mutáns allél elegendő a betegség (vagy tulajdonság) kifejeződéséhez.
AUTOSZOMÁLIS RECESSZÍV ÖRÖKSÉG- egy tulajdonság vagy betegség öröklődésének típusa, amelyben az autoszómában lokalizált mutáns allélt mindkét szülőtől örökölni kell.
AUTOSOME- minden kromoszóma, kivéve a nemi kromoszómákat; szomatikus sejtekben minden autoszóma kétszer van jelen. Az embernek 22 pár autoszómája van.
AFAKIA- a lencse veleszületett hiánya az ektoderma differenciálódási zavara miatt.
ACHEIRIA(apodia) - a kéz fejletlensége vagy hiánya.
KÉT VEGYÉRTÉKŰ- két konjugált homológ kromoszóma, amelyek mindegyike megkétszereződött. A profázis során megfigyelhető - I. meiózis.
CHORION BIOPSY- a terhesség 7-11. hetében végzett beavatkozás a születés előtti diagnózishoz szükséges sejtek kinyerésére.
BIOKÉMIAI SZŰRÉS ÖRÖKLETES METABOLIKUS HIBAJÁRA- újszülöttek vagy értelmi fogyatékosok kontingenseinek nem szelektív vizsgálata különféle biokémiai módszerekkel.
BLASTOPÁTIA- a blasztociszta károsodásából eredő fejlődési rendellenességek, pl. embrió a megtermékenyítés után legfeljebb 15 nappal.
BLEFAROPHIMOSIS- a szemhéjak vízszintes lerövidülése, i.e. a szem szűkítése.
BLEPHAROCHALASIA- a felső szemhéjak bőrsorvadása.
BRACHYDACTYLY- az ujjak lerövidülése.
BRAHIKAMPTODACTYLY- a kézközépcsont (meta-tarsalis) csontok és a középső phalangusok rövidülése camptodactyly-val kombinálva.
BRACHYCEPHALIA- a fej keresztirányú méretének növekedése a hosszanti méret relatív csökkenésével.
A klasszikus értelemben a gén egyszerre szolgált a genetikai rekombináció, mutáció és funkció egységeként. Később a genetikai lókuszt olyan kromoszómaszegmensként kezdték érteni, amelynek minden szerkezeti változása allélmutációs hatást vált ki, és amelyen belül keresztezés okozta törések is előfordulhatnak. A gén képes más anyagok reduplikációjára és szintézisére ("egy gén - egy polipeptidlánc").
VITILIGO- a bőr fokális depigmentációja.
veleszületett betegségek- születéskor elérhető.
A GENETIKAI KÓD DEGENERITÁSA- egy aminosav több kodonnak felel meg. A kodon harmadik bázisának cseréje nem mindig eredményez aminosavszubsztitúciót.
IVARSEJT- haploid kromoszómakészletet tartalmazó érett csírasejt.
GAMETOPÁTIA- veleszületett fejlődési rendellenességek, amelyek a csírasejtek (ivarsejtek) mutációján alapulnak.
HAPLOID KROMOSZÓMA KÉSZLET(P) - egy halmaz, amelyben minden kromoszóma egyszer van ábrázolva.
GAMARTOMA- az embrionális fejlődés megsértéséből származó daganatszerű képződés.
HEMERALÓPIA- a látás éles romlása gyenge fényviszonyok mellett, éjszakai vakság.
HEMIZIGOUSSÁG- a test állapota, amelyben egy gén jelen van az egyik kromoszómán, például a nemi kromoszómák nem homológ régióiban lévő gének.
GÉN- a DNS-ben található nukleotidszekvencia, amely meghatározott funkciót határoz meg: fehérjék vagy RNS szintézisét kódolja, vagy más gén transzkripcióját biztosítja. A "gén" kifejezést Johansen vezette be 1909-ben, mint egyfajta matematikai jellemzőt a valós, független, az ismeretlen természetű öröklődési egységeket kombináló és felosztó, keresztezési kísérletekben felfedezett, de bizonyos hatású öröklődési egységekre.
GÉNMANIPULÁCIÓ- technikák, módszerek és technológiák összessége rekombináns RNS és DNS kinyerésére, gének izolálására organizmusokból (sejtekből), gének manipulálására és más organizmusokba való bejuttatására.
GÉNTERÁPIA- genetikai anyag (DNS vagy RNS) bevitele egy sejtbe, amelynek funkciója megváltozik.
GENÓM- gének halmaza haploid sejtben.
GENOTÍPUS- a szervezet minden genetikai információja, amely fenotípusos kifejezéssel rendelkezik; egy szervezet genetikai jellemzői egy vagy több vizsgált lókuszban.
GÉNÁLLOMÁNY- egy adott populáció egyedeiben található allélkészlet.
A gének a modern adatok szerint a genetikai anyag lokalizált DNS-tartalmú régiói, amelyek specifikus funkciókban különböznek egymástól. Azokban a személyekben, akiknek sejtjei a sejtmagba és a citoplazmába differenciálódnak, a géneket a kromoszómák specifikus szakaszainak (lókuszainak) tekintik. A környezettel állandó kölcsönhatásban lévén G. befolyásolja a szervezet kialakulását és fejlődését; a gén károsodása működési zavarokhoz vezet.
HETEROSZIGÓTA Olyan organizmus, amely két különböző allélt tartalmaz a homológ kromoszómák adott helyén.
HETEROKROMATIN- egy kromoszóma vagy kromoszóma olyan szakasza, amely sűrű, tömör szerkezetű és genetikailag inaktív.
AZ ÍRISZ HETEROKROMIA- az írisz különböző részeinek egyenetlen színezése.
HIDROFTALMOUS(buphthalmos) - a szemgolyó növekedése.
HYDROCEPHALUS- a cerebrospinális folyadék túlzott felhalmozódása a koponyaüregben.
HIPERKERATÓZIS- az epidermisz stratum corneumának túlzott megvastagodása.
HIPERTELORIZMUS- a szervek közötti távolság növekedése (általában a szem körül).
HIPERTRICHOSIS- Túlzott szőrnövekedés.
SZELZELETEZETT HYPOPLASIA- egy szerv fejletlensége, amely a relatív tömegének vagy méretének hiányában nyilvánul meg.
HIPOSPADIA- a húgycső alsó hasadéka külső nyílásának elmozdulásával.
HIPOTELORIZMUS- a szervek közötti távolság csökkentése (általában a szem körül).
HIPOTHIROZIS- a pajzsmirigy elégtelensége. HIRSUTIZMUS- túlzott szőrnövekedés lányoknál férfi típus. HISTONES- a fő fehérjék, amelyek komplexeket képeznek a kromoszómában a DNS-sel. GLAUKÓMA- fokozott szemnyomás. HOLANDRI ÖRÖKSÉG- Y-kapcsolt öröklődés.
HOLOPROSENCEFALY- a telencephalon nincs félgömbökre osztva, és egy félgömb képviseli, egyetlen kamrai üreggel, amely szabadon kommunikál a subarachnoidális térrel.
HOMOZIGÓTA- olyan organizmus, amely két azonos allélt tartalmaz a homológ kromoszómák adott lokuszában.
HOMOLÓGIAI KROMOSZÓMÁK- méretben és alakban azonos kromoszómák, valamint az általuk hordozott gének számában és típusában. Ez alól kivételt képeznek a nemi kromoszómák.
KAPCSOLÓCSOPORT- az azonos kromoszómán található összes gén összessége.
DAKTILOSZKÓPIA- az ujjak bőrmintázatának tanulmányozása.
GÉNDISTINGULOSCOPIA- a tandem DNS ismétlődések számának és hosszának eltéréseinek kimutatása.
KALITONIZMUS Színlátási zavar, amelyre jellemző, hogy nem tud különbséget tenni a vörös és a zöld színek. Az öröklődés típusa X-hez kötött, recesszív.
DEXTROCARDIA- a szív helye a jobb oldalon.
TÖRLÉS- a kromoszómamutáció egy fajtája, amelyben a kromoszóma egy része elvész; a génmutáció olyan típusa, amelyben a DNS-molekula egy része hiányzik.
ELMEBAJ- szerzett demencia. A kognitív folyamatok szintjének tartós, visszafordíthatatlan csökkenése, érzelmek elszegényedése és viselkedési zavarok.
DIASTÉMA- rés a felső állkapocs központi metszőfogai között. Általában alacsonyan fekvő kantárral kombinálják.
Meckel-divertikulum- a vitelline csatorna intraabdominalis részének proximális szegmensének nem zárása; az ileum falának kiemelkedése.
DISCORIA- "macskaszem", a pupilla rés formájában.
DISTICHI A3- dupla sor szempilla.
DNS- Dezoxiribonukleinsav- biológiai makromolekula, genetikai információ hordozója.
ASE DNS POLIMER A DNS-szintézisben részt vevő enzimek.
DOLICHOCEPHALY- a fej hosszirányú méreteinek túlsúlya a keresztirányú méretekkel szemben.
DUPLIKÁCIÓ- a kromoszómamutáció olyan típusa, amelyben a kromoszóma bármely része megkétszereződik; egyfajta génmutáció, amelyben egy DNS-darab megkettőződik.
EUGENIKA- az emberi fejlesztés doktrínája genetikai módszerekkel.
A zigóta egy diploid sejt, amely egy petesejt és egy sperma fúziójából származik.
GENETIKAI PROBE – ismert szerkezetű vagy funkciójú DNS vagy RNS rövid darabja, amely valamilyen radioaktív vagy fluoreszcens vegyülettel van megjelölve.
IMMUNITÁS - a szervezet stabilitása (rezisztencia, rezisztencia, immunitás) különböző tényezőkkel szemben, amely lehetővé teszi saját integritásának és biológiai egyéniségének megőrzését.
A GENOMIKUS (genetikai vagy kromoszómális) imprinting olyan jelenség, melynek lényege a homológ kromoszómák (vagy gének) származásuktól (anyai vagy apai) függően eltérő aktivitása.
BELTÉTELT HÁZASSÁGOK - a rokonság második és további fokú vérrokonai között.
INVERZIÓ – a kromoszómális mutáció olyan típusa, amelyben a kromoszómák egy régiójában a gének sorrendje megfordul; a génmutáció olyan típusa, amelyben egy bázisszekvencia megfordul a DNS egy meghatározott régiójában.
INIONCEPHALIA - az occipitalis csont egy részének vagy egészének hiánya a foramen magnum kiterjedésével, aminek következtében az agy nagy része a hátsó koponyaüreg régiójában helyezkedik el.
BEHELYEZÉS - a génmutáció olyan típusa, amelyben a DNS egy szegmense beépül a gén szerkezetébe.
INTERFÁZIS – a sejtciklus sejtosztódások közötti fázisa, pre-szintetikus (G1), szintetikus (S) és posztszintetikus. (G2) időszakokban.
INTRON- az átírt DNS egy szakasza, amely a splicing során allokálódik, és nem vesz részt a polipeptidlánc kódolásában.
CAMPOMELIA - a végtagok görbülete.
KARIOTÍPUS - a fajra jellemző kromoszómakészlet (kromoszómák száma, mérete, alakja) sajátosságainak összessége, a kariotípus változása kromoszóma- és genomimutációk miatt következhet be.
KERATOKOPUS - elvékonyodott és heges szaruhártya kúpos kiemelkedése.
CLUSTER - a kromoszóma egy bizonyos régiójában elhelyezkedő különböző gének csoportja, amelyeket közös funkciók egyesítenek, például a hisztonfehérjék gének klasztere.
CELL CIKLUS - egyértelműen meghatározott események sorozata egy cellában, változó áramlási idejével.
CLINODACTYLY - az ujj oldalirányú vagy mediális görbülete.
A GÉNKLÓNOZÁS egy adott DNS szegmens millióinak azonos másolatainak előállítása mikroorganizmus felhasználásával erre a célra.
GENETIKAI KÓD - egyetlen rendszer az örökletes információk rögzítésére a DNS-ben.
KO DOMINÁNS ALLÉL - olyan, amely heterozigótában nyilvánul meg (például LV vércsoport).
KODOMINÁCIÓ - mindkét allél jeleinek megnyilvánulása heterozigóta egyedekben.
CODON (TRIPLET) - három nukleotidból álló szekvencia egy DNS-molekulában (vagy mRNS-ben), amely a fehérjemolekulában található aminosavak egyikét kódolja, vagy "pontozási jeleket" határoz meg az információk olvasásakor.
COLOBOMA - a szem résszerű hibája.
KIEGÉSZÍTŐ - a megfelelő bázisok szekvenciája a DNS ellentétes szálaiban.
COMPTODACTYLY - az ujjak proximális interphalangealis ízületeinek flexiós kontraktúrája.
KONJUGÁCIÓ - két homológ kromoszóma bivalenssé (tetradjává) való konvergencia és egyesülés, amelyek mindegyike megduplázódik, és a meiózis I. fázisában fordul elő.
Cordocentesis - vérvételi eljárás a magzat köldökvénájából.
CORRECTOPIA - a pupilla veleszületett elmozdulása.
Beletenyésztési együttható - annak a valószínűsége, hogy egy egyedben egy adott lókuszban két allél ugyanattól az őstől származik.
CRANIOSYNOSTOSIS - a koponyavarratok korai túlnövekedése, ami korlátozza a koponya növekedését és deformációjához vezet.
CRYPTOPHTALMOUS - a szemgolyó, a szemhéjak és a szemrepedés fejletlensége vagy hiánya.
CROSSINGOVER - helyek cseréje homológ kromatidák között a meiózis során.
LAGOPHTALMOUS - a szemhéjak nem teljes záródása.
LEUKOCORIA - fehér pupilla.
LENTICONUS - a lencse egy részének kiemelkedése.
LISSENCEPHALIA - lásd agyria.
A LOCUS egy gén helye a kromoszómán.
MACROGLOSSIA - a nyelv kóros megnagyobbodása.
MACRODACTYLY - Az ujjak és/vagy lábujjak túlzott megnagyobbodása.
MACROCORN A - a szaruhártya átmérőjének növekedése.
MAKROSZÓMIA (gigantizmus) - a testméret túlzott növekedése.
MAKROSZTÓMIA - túlságosan széles szájrés.
Macrotia - megnagyobbodott fülkagyló.
MACROCEPHALIA - a koponya méretének növekedése.
MEGALOCORNEA - lásd makroszaru.
Megaloureter - az ureter kiterjesztése és meghosszabbítása.
MEIOSIS - egy éretlen csírasejt magjának két egymást követő (I és II) osztódása egy replikációs ciklus alatt, amelynek eredményeként érett csírasejtek képződnek haploid ivarsejt kromoszómakészlettel. A sejtosztódás módszere.
METAFÁZIS - a mitózis és a meiózis stádiuma, amelyben a kromoszómák orsó formájában sorakoznak fel az egyenlítőn, és metafázis lemezt alkotnak.
MICROGENIA - az alsó állkapocs kis mérete.
MIKROGNATIA - a felső állkapocs kis mérete.
MICROPOLYGYRIA - az agyféltekék kisszámú és rendellenesen elhelyezkedő konvolúciója.
MIKROSZTÓMIA - kis szájrés.
MICROTIA - a fülek méretének csökkenése.
MICROPHAKIA - a lencse kis mérete.
MIKROFTALMIA - a szemgolyó kis mérete.
MICROCEPHALIA - az agy és az agykoponya kis mérete.
MICROCORN A - a szaruhártya átmérőjének csökkenése.
A MISSENS MUTÁCIÓK olyan génmutációk, amelyek az egyik aminosav másik aminosavval való helyettesítését eredményezik.
A MITOSIS a sejtosztódás egy fajtája, amelyben a leánymagok ugyanannyi kromoszómát hordoznak, mint a szülősejt. A szomatikus sejtek osztódásának módja.
MITOKONDRIÁLIS ÖRÖKSÉG – a mitokondriális DNS-en keresztül továbbított tulajdonságok öröklődése.
TÖBB ALLÉL - ugyanazon lókusz kettőnél több alléljának jelenléte.
MÓDOSÍTÁS - fenotípusos, nem örökletes változások, amelyek a hatása alatt következnek be különféle tényezők környezet.
MOZAIK – olyan egyén, akinek különböző kromoszómakészletű sejtjei vannak.
MOZAICIZMUS - a kromoszómakészlet heterogenitása a test különböző sejtjeiben. Egyes sejtek kromoszómakészlete rendellenes, míg mások normálisak lehetnek. A teljes kromoszómális anomáliától (gametikus) ellentétben a rendellenesség a zigóta hasításának korai szakaszában jelentkezik.
MONGOLOID SZEMSZAKCIÓ - a palpebrális repedések belső sarkai le vannak süllyesztve.
MONOAPUS - egy alsó végtag hiánya.
MONOBRACHY - egy felső végtag hiánya.
MONODAKTILIA - egy ujj jelenléte a kézen vagy a lábon, az oligodaktilia egy fajtája.
MONOSOMIKUS - olyan sejt, szövet, szervezet, amelynek kromoszómakészletéből az egyik kromoszóma hiányzik.
MONOSZÓMIA - az egyik kromoszóma hiánya a diploid halmazban.
A GYŰJTETETT MORFOGENETIKAI VÁLTOZATOK (syn.: fejlődési mikroanomáliák, jelek vagy stigmák, a dysembryogenesis) olyan fejlődési eltérések, amelyek túlmutatnak a normál eltéréseken, de nem sértik a szerv funkcióit.
Az mRNS - az eukarióták fehérjeszintézisének templátja, a sejtmagban feldolgozás és splicing eredményeként képződik, és átjut a citoplazmába, ahol az információ lefordításra kerül.
A mukopoliszacharidózis a mukopoliszacharidok metabolizmusának veleszületett rendellenessége.
MULTIFAKKURIÁLIS BETEGSÉGEK - azok, amelyek a különböző lókuszok alléljainak bizonyos kombinációinak és a környezeti tényezők specifikus hatásainak kölcsönhatása következtében alakulnak ki.
MUTAGÉN – mutációt okozó tényező.
MUTÁNS – mutáns allélt hordozó organizmus.
FRAME HIFTING MUTATIONS - a DNS-molekula olyan szakaszainak deléciója vagy inszerciója (inszerciója), amelyek mérete nem többszöröse három bázisnak.
MUTÁCIÓ - az örökletes struktúrák (gén, kromoszóma, genom) változása.
Nanizmus – törpeség.
"CAPE WIDOW" - ék alakú szőrnövekedés a homlokon.
ÖRÖKLETES BETEGSÉG - olyan betegség, amelynek etnológiai tényezője gén, kromoszóma vagy genomi mutáció.
ÖRÖKLETESSÉG – a genetikai tényezők miatti általános fenotípusos variabilitás része.
NEVUS - anyajegy, anyajegy: korlátozott bőrdiszplázia folt vagy daganatszerű képződmény formájában.
FOLYAMATOS VÁLTOZATOSSÁG – ez a fajta változékonyság, amelyben az egyedek nem tartoznak egyértelműen meghatározott fenotípusos osztályokba; mennyiségi tulajdonságokra jellemző.
A KROMOSZÓMÁK NEM DISZPERZIÓJA - jelenség a sejtosztódás során: ennek eredményeként mindkét homológ kromoszóma vagy testvérkromatidák ugyanarra a pólusra költöznek, és aneuploid sejteket képeznek.
NYSTAGMUS – önkéntelen, gyorsan követi a szemmozgásokat egyik oldalról a másikra (néha körkörös vagy felfelé és lefelé), a központi idegrendszer betegségével kapcsolatos.
NONSENSE MUTÁCIÓK - génmutációk, amelyek szensz kodon helyett terminátor kodon kialakulásához vezetnek.
REACTION RATE - a környezeti hatásokra való reagálás sajátos módja, amelyet a genotípus jellege határoz meg.
NUKLEOSZÓMA – egy kétszálú DNS-ből (körülbelül 200 bázispárból) álló kromoszóma szegmense, amely egy fehérjemag-mag köré van tekercselve, és hiszton fehérjemolekulákból áll.
Nukleotid - DNS vagy RNS monomer, amely nitrogéntartalmú bázisokat, szénhidrátot és foszforsav maradékot tartalmaz.
A NULLISOMIC egy aneuploid, amelynek kariotípusából hiányzik egy pár homológ kromoszóma.
OXYCEPHALIA – lásd acrocephalia.
OMFALOCELE - a köldökzsinór sérve.
ONTOGENEZIS – a szervezet egyéni fejlődése.
Az OOGENESIS a sejtek differenciálódásának és érésének folyamata, amely a női ivarsejtek (peték) kialakulásához vezet.
Az OOGONIUM egy primordiális csírasejt, amely mitózisban petesejteket hoz létre.
OOCYT - női csírasejt, amelyből a meiózis és az érés eredményeként petesejt képződik (P).
TERMÉKEZÉS - az ivarsejtek (pete és hímivarsejtek) összeolvadása a képződéssel (2ga), amelyből többsejtű szervezet fejlődik ki.
SZERVEZET - az ontogenezis azon fázisa, amely során az embrionális szervek elkülönülnek a csírarétegektől és megkülönböztetik őket.
ORTHODACTYLY - lásd symphalangia.
Pachygyria - a fő kanyarulatok megvastagodása.
Pachyonychia - a körmök megvastagodása.
PENETRANCE - egy tulajdonság megnyilvánulásának valószínűsége (vagy gyakorisága) a hordozókban domináns gén vagy homozigóta egyedekben recesszív gén.
PEROMELIA - rövid végtaghossz normál méretek torzó.
PILONIDAL fossa (sacral sinus, epithelialis coccygealis járat) - többrétegű csatorna laphám, a farkcsontnál az intergluteális redőben nyíló.
Platibasia - a koponyaalap ellaposodása, amelyet általában a foramen magnum deformációja kísér.
Platyspondylia - a csigolyák ellaposodása.
PLEIOTROPIA - egy gén hatásának megnyilvánulásainak sokfélesége különböző szervekben és/vagy szövetekben.
Abszolút lokalizációjuk szerint a gének fel vannak osztva autoszomális és nemhez kötött. A nemi kromoszómában egy gén lokalizálható az X kromoszóma egy olyan szegmensében, amely nem homológ az Y kromoszómával (abszolút X-kötés), az Y kromoszóma olyan szegmensében, amely nem homológ az X kromoszómával (holandrikus gének abszolút Y-kötés), vagy homológ X és Y kromoszómák szegmensén (nem teljes nemi kapcsolat). Relatív lokalizációjuk szerint bármely két nem allél gén tartozhat ugyanabba vagy különböző kapcsolódási csoportba (kromoszómákba).
POLIGÉN JELLEMZŐK- sok gén által okozott tulajdonságok, amelyek mindegyike csak kis mértékben befolyásolja ennek a tulajdonságnak a kifejeződési fokát.
POLYDACTYLY- az ujjak számának növekedése.
A KORLÁTOZÁSI SZEDÉKEK HOSSZÁNAK POLIMORFIZMÁJA (RFLP)- DNS-szegmensek jelenléte különböző hosszúságú feldolgozás után DNS bizonyos korlátozások.
POLIPEPTID- a peptidkötéssel összekapcsolt aminosavak szekvenciája.
POLIPLOID Olyan szervezet, amely három vagy több kromoszómakészlettel rendelkezik.
POLIPLOIDIA- olyan jelenség, amely a kromoszómák számának megváltozásához vezet a test sejtjeiben, a haploid többszöröse.
POLYSOMA- mRNS-molekulák komplexe, amelyen több aktív riboszóma található, amelyek mindegyike egy-egy fehérjemolekulát szintetizál.
Udvarias LIA- Túl sok mellbimbó.
SZEX KROMATIN- festőtest (inaktivált X kromoszóma) a sejtmagban, amelynek száma mindig eggyel kevesebb, mint az X kromoszómák száma.
SEX KROMOSÓMÁK- kromoszómák, amelyek a két nem között különböznek, és X-nek és Y-nek vannak jelölve.
PORENCEFALIA- ependimával bélelt üregek jelenléte az agyszövetben, amelyek az agy kamráival és a subarachnoidális térrel kommunikálnak.
PREAURICULAR PAPILLOMÁK- a külső fül töredékei, amelyek a fülkagyló előtt helyezkednek el.
PREAURIKULIS SIGYLÁK(preauricularis gödrök) - vakon végződő járatok, amelyek külső nyílása a fülkagyló felmenő részének alján található a tragus vagy lebeny előtt.
GENETIKAI HASZNÁLAT- különböző lókuszok mutáns alléljei vagy allélkombinációi, amelyek hajlamosak a környezeti tényezők hatására kialakuló betegségekre és azok súlyosabb lefolyására.
PRENATAL DIAGNÓZIS- A magzati fejlődés során fellépő örökletes betegségek vagy egyéb rendellenességek diagnosztizálása.
PROBAND- az a személy, akire vonatkozóan a törzskönyv épül.
ORMÁNY- törzs alakú nyúlvány, amely az orr gyökerénél található, vagy egyetlen szemrepedés felett ciklopiával.
PROGENIA- Az alsó állkapocs előrenyúlása a felsőhöz képest.
PROGERIA- korai öregedés.
PROGNATIA- A felső állkapocs előrenyúlása az alsóhoz képest.
PROSENCEFALY- az elülső agyhólyag elégtelen felosztása nagy féltekékre.
VIZSGÁLATI PROGRAMOK(lásd szűrés).
pterygium- pterigoid bőrredők.
PTOSIS- kihagyás (általában szemhéj).
OLVASÓKERET- a nukleotidszekvencia szekvenciális hármassorok formájában történő leolvasásának három lehetséges módja egyike.
HASÍTOTT- egymástól eltérő egyedek megjelenése az utódokban (különböző fenotípusos és genotípusos osztályok).
REKOMBINÁCIÓ- a gének átrendeződése a hibridekben az ivarsejtek kialakulása során, valamint a szülők összetételének átrendeződése. Ezek a jelenségek a tulajdonságok új kombinációihoz vezetnek az utódokban.
JAVÍTÁS- a sérült DNS szerkezet helyreállítása.
REPLIKÁCIÓS VILLA (REP L IKON)- a DNS-molekula azon része, amelyben az új DNS szintézise egyszálú formában történik.
DNS REPLIKÁCIÓ- az a folyamat, amelynek során az anya-DNS-molekulák bázisszekvenciájában kódolt információ maximális pontossággal kerül továbbításra a leány DNS-be.
KORLÁTOZÁSOK- bizonyos bázisszekvenciákra erősen specifikus enzimek, amelyek a molekulát hasítják DNS.
RECESSZÍV GÉN- egy ilyen gén, amelynek expresszióját e gén többi allélja elnyomja, csak homozigóta állapotban jelenik meg.
RIBOSZÓMA- a citoplazma organoidja, amely nagy és kis szubrészecskékből áll, és amelyen a polipeptid szintézise történik.
RIZOMELIA- a proximális végtagokhoz kapcsolódik.
RNS- ribonukleinsavak.- nukleinsavak egyszálú polimer molekulái, amelyek részt vesznek a fehérje bioszintézis folyamataiban és különböző funkciókat látnak el (mRNS, tRNS, rRNS, hnRNS).
CSALÁDFA- egy diagram, amely egyazon család tagjainak kapcsolatát mutatja be generációk sorozatában.
rRNS- RNS, amely fehérjével komplexben riboszómát alkot, amelyben fehérjeszintézis megy végbe.
WEBOLDAL- egy nukleinsav molekula része.
SZINUSSZINSZ- lásd pilonidal fossa.
SZEKVENCIÁZÁS- a DNS nukleotid szekvencia meghatározása restrikciós enzimek segítségével.
CSALÁDI BETEGSÉGEK- több családtagnál megfigyelt betegségek egy vagy több generációban.
SIBS- testvérek.
SYMPHALANGIA(orthodactyly) - az ujj falángjainak fúziója.
SYNDACTYLY- a szomszédos ujjak vagy lábujjak teljes vagy részleges összeolvadása.
GENETIKUS SZINDRÓMA- sok egyidejűleg előforduló tulajdonság genetikailag szabályozott komplexe, amely gyakran egy gén pleiotróp (többszörös) hatásához kapcsolódik.
SYNECHIA- a szomszédos szervek felületét összekötő rostos szálak.
SZINOSTÓZIS- a csontok nem szétválása (fúziója).
SINOFRIES- lógó szemöldök.
SIRENOMELIA- az alsó végtagok összeolvadása.
Scaphocephaly- az idő előtt túlnőtt sagittalis varrat helyén kiálló taréjú, megnyúlt koponya.
SKLEROKORNÉA- a szaruhártya diffúz homályosodása, amelyben a szaruhártya fehér, nehezen megkülönböztethető a sclerától.
SZŰRÉS (SZŰRÉS)- különböző kontingensek tömeges vizsgálata a patológiás személyek előzetes azonosítására.
toldás- az intronok eltávolításának és az exonok érett mRNS-vé való kombinálásának folyamata.
SZŰKÜLET- a csatorna vagy nyílás szűkítése.
LÁBHINTÁZÓ SZÉK- megereszkedett ívű láb, kiálló sarokkal.
SZTRABIZMUS- strabismus.
SZFEROFAKIA- gömb alakú lencse.
GÉNÖSSZETÉTEL- gének (jellemzők) együttes átvitele, a gének ugyanabban a kromoszómában történő lokalizációja miatt.
TANDEM SZEKVENCIÁK- ugyanazon szekvenciák több másolata, egymás után, azonos irányba orientálva.
TAURODONTIZMUS- a fog üregének jelentős növekedése.
TELEANGIECTASIA- a kapillárisok és a kiserek helyi tágulása.
TELEKANT- a palpebralis repedések belső sarkainak oldalirányú elmozdulása normál elhelyezkedő pályákkal.
BIKA BARR- szexkromatin.
MEGSZŰNÉS- a folyamat vége.
ÁTÍRÁS- örökletes információk olvasása génexpresszió során; genetikai információ átvitele a DNS-ből a RNS.
FORDÍTÁS- kromoszómamutáció, amelyet egy kromoszómaszegmens helyzetének megváltozása jellemez.
ADÁS- örökletes információk továbbítása; fehérje molekula szintézise vagy az mRNS bázisszekvencia transzlációja a polipeptid lánc aminosavszekvenciájává.
TRIGONOCEFALIA- a koponya kitágulása az occipitalisban és szűkület az elülső részben.
"LÓHERE"- a koponya rendellenes formája, magasan kiálló homlok, lapos nyakszirti, a halántékcsontok kiemelkedése jellemezhető, a parietálishoz kapcsolva mély mélyedéseket határoznak meg.
TRIRADIUS- a bőrmintázat egy eleme - három gerinc konvergenciájának helye.
triszómia- egy további kromoszóma jelenléte egy diploid szervezet kariotípusában; a poliszómia olyan típusa, amelyben három homológ kromoszóma van (a triszómiában szenvedő egyedet triszómiának nevezik).
tRNS- specifikus aminosav-maradékokat hordozó RNS átvitele az mRNS meghatározott területére.
SZÍJAK SIMON AR A- lásd a magzatvíz sávokat.
Fenilketonúria- a szervezetben a fenilalanin metabolizmusának megsértése genetikai hiba miatt. A születés utáni első hónapokban található meg, a betegek leggyakrabban az imbecilitás és az idiotizmus szintjét érik el. Az öröklődés típusa autoszomális recesszív.
FENOKÓPIA- a fenotípus nem örökletes változása, amely hasonló bizonyos mutációk megnyilvánulásához.
FENOTÍPUS- egy szervezet megfigyelt külső és belső jeleinek összessége az ontogenezis adott szakaszában, amely a genotípus és a külső környezet kölcsönhatásának eredményeként jön létre.
FETOPÁTIA- a magzat károsodása a 9. héttől a szülésig. FETOSKÓPIA- olyan eljárás, amely lehetővé teszi a magzat méhen belüli vizsgálatát száloptikai technológia segítségével.
MAGZATTERÁPIA (MAGZATTERÁPIA; SZÜLETÉS ELŐTT TERÁPIA)- a magzat szülés előtti kezelése.
SZŰRŐ- az alsó orrpont és a felső ajak vörös határa közötti távolság.
PHOCOMELIA- a proximális végtagok hiánya vagy jelentős fejletlensége, aminek következtében a lábak vagy kezek közvetlenül a testhez tapadnak.
CHOREA HUNTINGTON- betegség, amelynek fő jellemzői: chorea (a célzott cselekvés képtelensége, a végtagok hiperkinéziája) és demencia. Az öröklődés típusa autoszomális domináns, magas penetranciával.
KROMATIDOK- a reduplikált kromoszóma alegységei, jövőbeli kromoszómák.
KROMATIN- DNS-molekula komplexben hisgon fehérjékkel. A spiralizáció következtében a DNS mérete csökken, ami kromoszóma kialakulásához vezet.
KROMOSZOMÁLIS MUTÁCIÓ (VAGY ABERRÁCIÓ)- a kromoszómák szerkezetének megváltozása.
KROMOSZOMÁLIS KÉSZLET- kromoszómakészlet egy normál ivarsejt vagy zigóta magjában.
KROMOSZÓMÁK- a sejtosztódás során látható sejtmag szuborganoidjai, amelyek bizonyos alakúak és szerkezetűek, nagyszámú gént tartalmaznak, és képesek önreprodukcióra.
X-LINKEDÖRÖKSÉG – azon tulajdonságok öröklődésének típusa, amelyek génjei az X kromoszómán találhatók. t CEBOCEPHALIA- "majom" fej.
CENTROMÉREK- a kromoszóma azon régiója, amelyhez a szálak kapcsolódnak
orsó a sejtosztódás során.
CIKLÓPIA- egyetlen szem jelenléte egy pályán, a homlok középvonala mentén.
EXONUCLASES- olyan enzimek, amelyek szekvenciálisan hasítják le a nukleotidokat a DNS 5", 3" vagy RNS.
EXONS- egy nem folytonos gén fragmentumai, amelyek információt hordoznak a polipeptid aminosav-szekvenciájáról, intronokkal megszakítva és az érett mRNS-ben megőrződnek.
EXOFTALMOUS- a szemgolyó előretolódása, a palpebrális repedés tágulása kíséretében.
EXENCEPHALIA- a koponyaboltozat csontjainak hiánya és a fej lágy szövetei, aminek következtében az agyféltekék nyíltan helyezkednek el a koponyaalapon, különálló csomópontok formájában, amelyeket pia mater borít.
KIFEJEZÉS- egy genetikailag meghatározott tulajdonság kifejeződési foka.
GÉNKIFEJEZÉS- a géntranszkripció aktiválása, melynek során mRNS képződik a DNS-en.
HÓLYAGEXTRÓFIA- veleszületett hólyag- és hasfalhasadék kiemelkedéssel hátsó fal hólyag a hasizmok hibáján keresztül kifelé.
ECTOPIA- a szerv elmozdulása, i.e. szokatlan helyen található.
A LENCE ECTOPIA- a lencse elmozdulása az üvegtestből.
EKTRODAKTÍLIA- a kéz (láb) központi komponenseinek apláziája "karomszerű" kéz (láb) kialakulásával.
EMBRYOPÁTIA- a megtermékenyítést követő 16. naptól a 8. hét végéig tartó időszakban jelentkező hiba.
EMBRYOTOXON hátsó- a szaruhártya perifériás homályosodása gyűrű vagy félgyűrű formájában.
ENDONUCLEASES- a polinukleotid láncon belüli kötéseket hasító enzimek bizonyos nukleotidszekvenciákra specifikusak.
ENTROPION- a szemhéj hibája, amelyben a szemhéj széle a szemgolyó felé tekered, ami a szaruhártya károsodásához vezet a szempillák által.
EPIBULBAR DERMOID- lipodermoid növedékek a szemgolyó felszínén, gyakrabban az írisz és a sclera határán.
EPIKANT- a belső cantusz függőleges bőrredője.
EPISPADIUM- felső húgycső hasadék, amelyet a pénisz görbülete kísér.
hámfossa- lásd pilonidal fossa.
ERITROBLASTÓZIS (ÚJSZÜLÜLETEK HEMOLITIKUS BETEGSÉGE)- a magzat és az újszülött betegsége, amelyet az Rh faktor szerinti immunkonfliktus, ritkábban az ABO rendszer vércsoportja okoz. Abban az esetben, ha egy Rh-negatív anyában Rh-pozitív magzat vagy egy O (I) vércsoportú anya fejlődik ki, ha a magzat más vércsoportú, különösen A (II).
EUKARIÓTÁK olyan élőlények, amelyek sejtjei membránnal körülvett maggal rendelkeznek.
EUKROMATIN- a kromoszómák azon szakaszai, amelyek az interfázisú magokban dekondenzáción mennek keresztül, funkcionálisan aktív genetikai anyagból állnak.
MAG- eukarióta sejtek létfontosságú organellumja, amelynek jellemzője a genetikai anyag (DNS) jelenléte.
NUCLELLAR SZERVEZŐ a kromoszóma azon régiója, amely tartalmazza a kódoló géneket rRNS.
TOJÁS- nőstényekben a gametogenezis során képződő csírasejt.
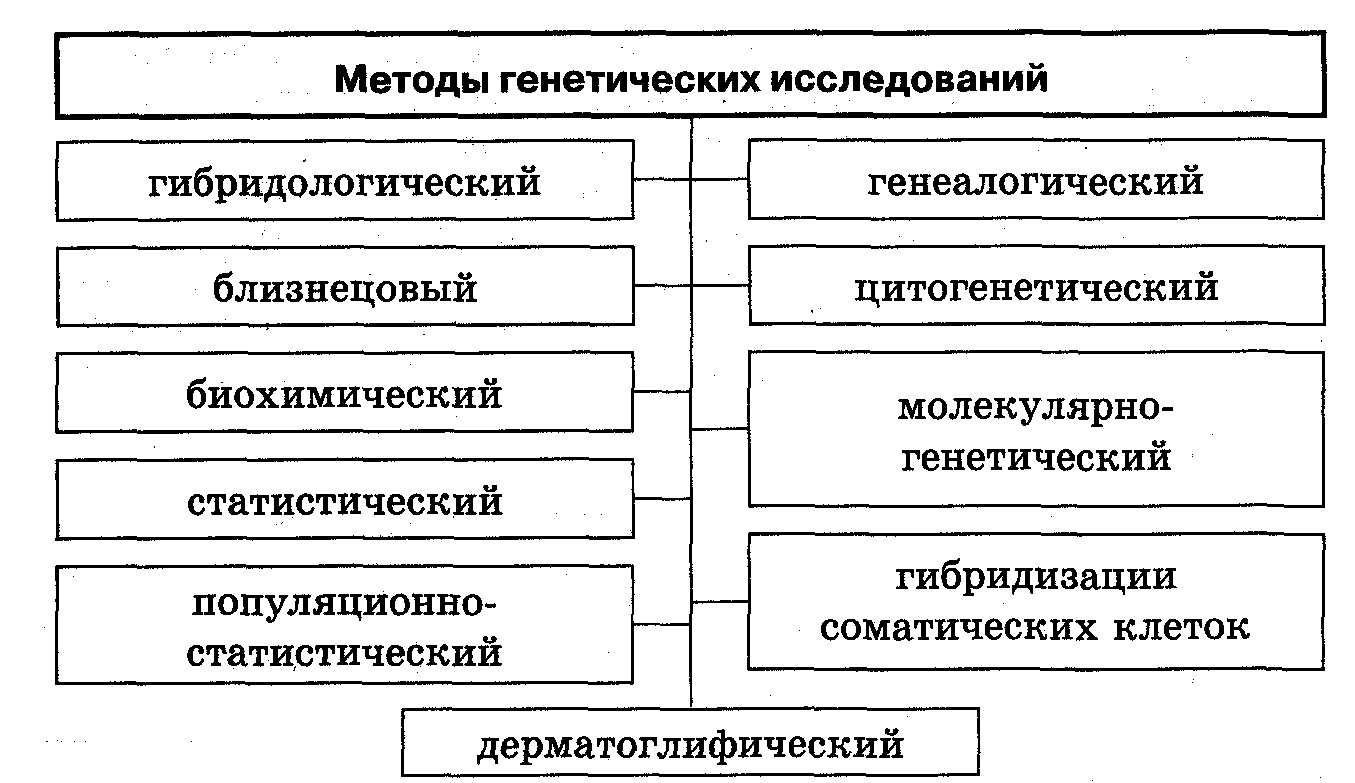

3.5. Az öröklődés mintái, citológiai alapjaik. G. Mendel által megállapított öröklődési minták, ezek citológiai alapjai (mono-hibrid keresztezés). T. Morgan törvényei: a tulajdonságok kapcsolt öröklődése, a gének kapcsolódásának megsértése. Szexgenetika. A nemhez kötött tulajdonságok öröklődése. A gének kölcsönhatása. Genotípus mint komplett rendszer. Az emberi genetika. Az emberi genetika tanulmányozásának módszerei


Kapcsolt öröklés- ugyanazon a kromoszómán található gének együttes öröklődése. A tengelykapcsoló jelenséget T. Morgan tanulmányozta. Kapcsolt öröklődés esetén megfigyelhető az átkelés jelensége - a homológ kromoszómák keresztezése a meiózis folyamatában és a kromoszómák közötti helyek cseréje

homogametikus szex- a nemi kromoszómán azonos ivarsejtek alkotják az ivart. Emberekben, emlősökben, Drosophilában a nőstény nem homogametikus, a pillangóknál, hüllőknél és madaraknál a férfi nem.
Heterogametikus szex- a nemi kromoszómán nem egyforma ivarsejtek által alkotott nem
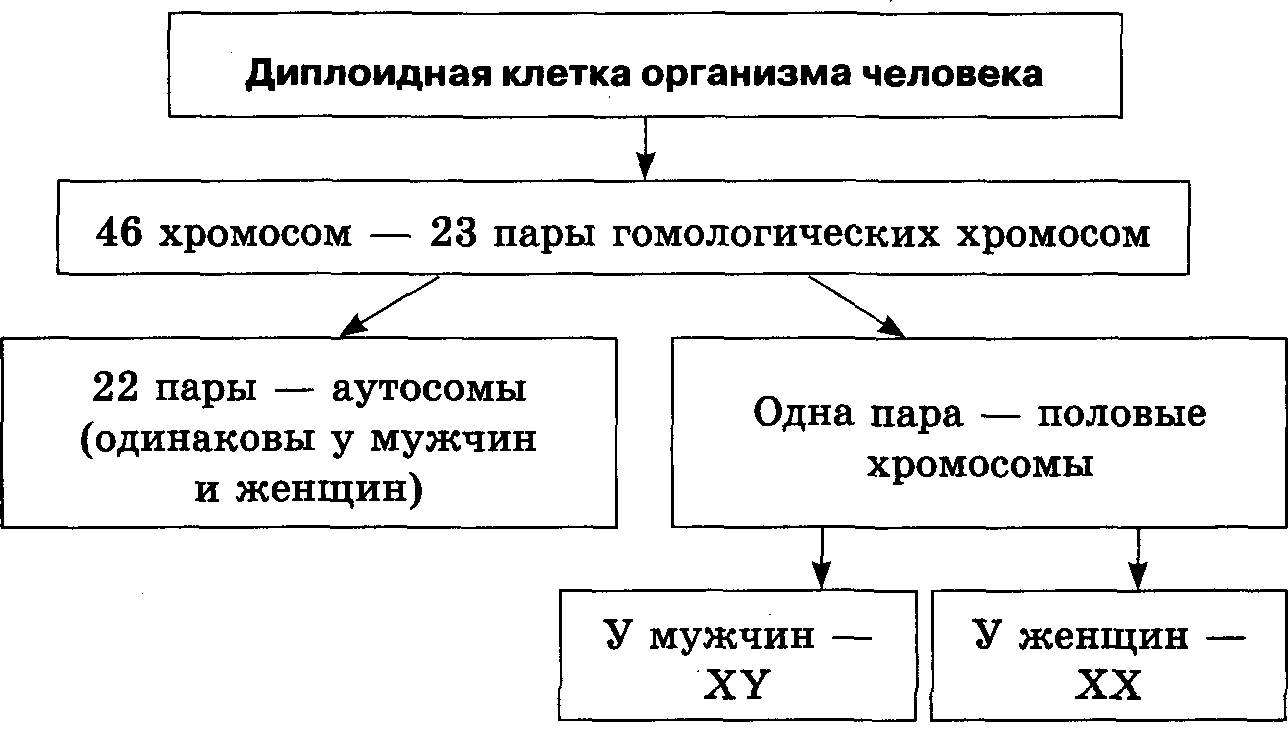
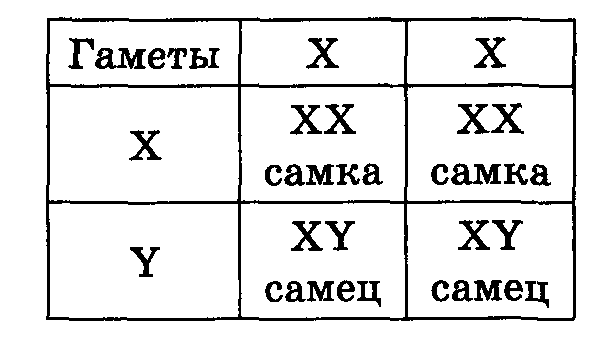
A szervezet nemét a fogantatás időpontjában határozzák meg; annak a valószínűsége, hogy hím vagy nőstény lesz, 1:1
A nemi kromoszómákon találhatók egyes gének, amelyek nem azonosak a különböző nemű egyedekben, így az általuk kódolt tulajdonságok öröklődésének megvannak a maga sajátosságai: az anya nemhez kötődő tulajdonságai már az első generációban megjelennek a fiakban. , és az apák vonásai a lányokban. Ha bármilyen tulajdonságot kódol az Y-kromoszóma gén, akkor azt csak a férfi vonalon keresztül továbbítják.
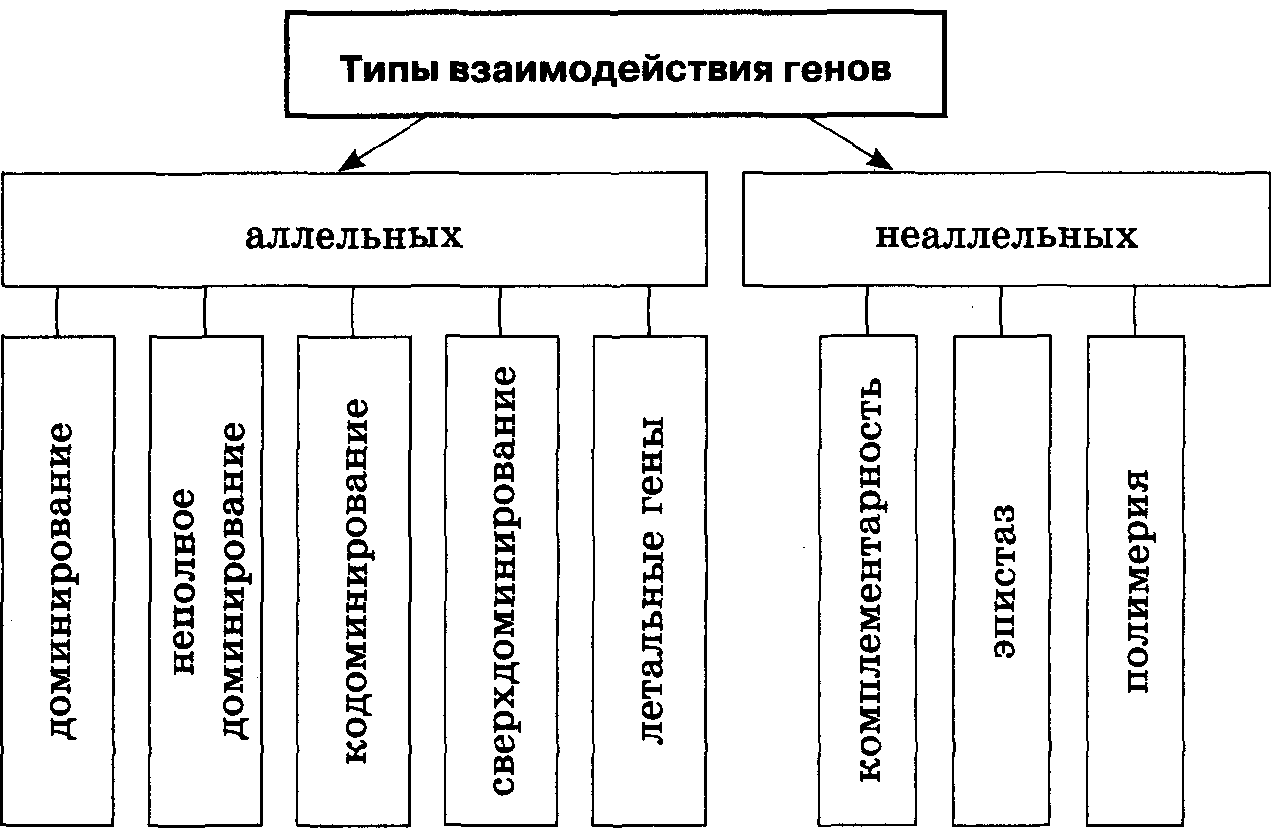
Genotípus- egy szervezet kölcsönható génjeinek integrált rendszere. A gének öröklődnek egymástól függetlenül, vagy szoros kölcsönhatásban állnak egymással. A genotípus integritása a faj evolúciója során alakult ki
Az öröklődés törvényei nemcsak a növényekre és állatokra vonatkoznak, hanem az emberre is. A szülőktől a gyermek olyan génkészletet kap, amely meghatározza az összes tulajdonság kialakulását. Az emberek, mint más szervezetek, domináns és recesszív tulajdonságokkal rendelkeznek.

vad típusú allél(normál) - egy gén nukleotidszekvenciája, amely biztosítja annak normális működését.
Allél domináns- allél, amelynek jelenléte a fenotípusban nyilvánul meg.
Allél mutáns- olyan mutáció, amely a vad típusú allél szekvenciájának megváltozásához vezet.
Recesszív allél- olyan allél, amely fenotípusosan csak homozigóta állapotban nyilvánul meg, és domináns allél jelenlétében takar.
kromoszóma aberráció(vagy kromoszóma-rendellenesség) a kromoszómamutációk bármely típusának általánosított neve: deléciók, transzlokációk, inverziók, duplikációk. Néha genomi mutációk (aneuploidiák, triszómiák stb.) is jelezhetők.
allél- egy gén két vagy több alternatív formája közül az egyik, amelyek mindegyikét egyedi nukleotidszekvencia jellemez; az allélek általában nukleotidszekvenciában különböznek egymástól.
Allél sorozat- monogén örökletes betegségek ugyanazon gén különböző mutációi okozzák, de klinikai megnyilvánulásaik szerint különböző nozológiai csoportokba tartoznak.
Amplicon- extrakromoszómális amplifikációs egység.
DNS-erősítő(thermocycler) - a polimeráz láncreakció (PCR) végrehajtásához szükséges eszköz; lehetővé teszi a kívánt ciklusszám beállítását és az optimális idő- és hőmérséklet-paraméterek kiválasztását minden egyes ciklusfolyamathoz.
Erősítés- a gének másolatainak számának növekedése (a DNS mennyisége).
ErősítésDNS- a DNS egy bizonyos szakaszának szelektív másolása.
Amfidiploidok- két genom egyesülésének eredményeként két kettős kromoszómakészletet tartalmazó eukarióta sejtek.
Aneuploidia- megváltozott kromoszómakészlet, amelyben a szokásos készletből egy vagy több kromoszóma hiányzik, vagy további másolatok képviselik.
Antikodon- egy három nukleotidból álló szekvencia a transzfer RNS-molekulában, amely komplementer az mRNS-molekulában található kódoló triplettel.
Antimutagenezis- az a folyamat, amely megakadályozza egy mutáció rögzülését (lehetőségét), azaz az eredetileg sérült kromoszóma vagy gén eredeti állapotába való visszaállítását.
Autosome bármely nem nemi kromoszóma. Az embernek 22 pár autoszómája van.
Autoszomális domináns öröklődés- az öröklődés olyan típusa, amelyben egy, az autoszómában lokalizált mutáns allél elegendő a betegség (vagy tulajdonság) kifejeződéséhez.
Autoszomális recesszív öröklődés- egy tulajdonság vagy betegség öröklődésének típusa, amelyben az autoszómában lokalizált mutáns allélt mindkét szülőtől örökölni kell.
bakteriofág- bakteriális vírus: fehérjehéjba csomagolt DNS-ből vagy RNS-ből áll.
Génbank (könyvtár).- egy adott szervezet génjeinek teljes készlete, amelyet a rekombináns DNS részeként nyernek.
fehérjemérnökség- kívánt tulajdonságokkal rendelkező mesterséges fehérjék létrehozása gének irányított változtatásával (mutációjával), vagy heterológ gének közötti lókuszok cseréjével.
Korion biopszia- a terhesség 7-11. hetében végzett beavatkozás a születés előtti diagnózishoz szükséges sejtek kinyerésére.
Southern blotting- módszer a DNS-próbával komplementer szekvenciákat tartalmazó DNS-szegmensek azonosítására szilárd mátrixon (nitrocellulóz vagy nejlon szűrők) rögzített elektroforetikusan elválasztott DNS-fragmensek között.
Blotting- DNS, RNS vagy fehérje molekulák átvitele a gélből, amelyben elektroforézis történt, a szűrőre (membránra).
Vakcina- gyengített vagy elpusztult fertőző ágens (vírus, baktérium stb.) vagy annak egyedi összetevőiből álló készítmény, amely antigéndeterminánsokat hordoz, és képes immunitást kiváltani ezzel a fertőzéssel szemben állatokban (emberekben). Ezenkívül a közelmúltban génmanipulált vakcinák váltak elérhetővé (ilyen vakcina például a hepatitis B vakcina).
Vektor- idegen DNS beépítésére és autonóm replikációra képes DNS-molekula, amely a genetikai információ sejtbe való bejuttatásának eszközeként szolgál.
Vektor a klónozáshoz- bármilyen kisméretű plazmidot, fágot vagy DNS-t tartalmazó állati vírust, amelybe idegen DNS beilleszthető.
Vírusok- nem sejtes természetű fertőző ágensek, amelyek képesek a sejtek anyagcseréjét a genomjukban kódolt genetikai információ megvalósítása során a vírusrészecskék szintézise felé irányítani. A vírusoknak lehet fehérjehéja, vagy csak DNS-ből vagy RNS-ből állhatnak.
veleszületett betegségek- a születéskor jelenlévő betegségek lehetnek örökletesek és a szervezet egyedfejlődési hibái is.
β-galaktozidáz- egy enzim, amely hidrolizál - β-galaktozidokat, különösen laktózt, szabad galaktóz képződésével.
Ivarsejt- Érett nemi sejt.
Haploid- egyetlen gén- vagy kromoszómakészletet tartalmazó sejt.
Hemizigótaság Egy szervezet állapota, amelyben egy gén egy kromoszómán található.
Gén- nukleotidszekvencia a DNS-ben, amely egy specifikus RNS-t kódol.
genetikai térkép- a szerkezeti gének és szabályozó elemek kromoszómában való elrendeződésének diagramja.
genetikai kód- megfelelés a DNS-ben (vagy RNS-ben) található tripletek és a fehérjék aminosavai között.
Génmanipuláció- technikák, módszerek és technológiák összessége rekombináns RNS és DNS kinyerésére, gének izolálására egy szervezetből (sejtekből), gének manipulálására és más organizmusokba való bejuttatására.
Génterápia- genetikai anyag (DNS vagy RNS) bejuttatása a sejtbe a normális működés helyreállítása érdekében.
Genom- egy szervezet génjeiben található általános genetikai információ, vagy egy sejt genetikai felépítése.
Genotípus 1) egy szervezet összes genetikai információja; 2) a szervezet genetikai jellemzői egy vagy több vizsgált lókuszra vonatkozóan.
Szabályozó gén- egy szabályozó fehérjét kódoló gén, amely aktiválja vagy elnyomja más gének transzkripcióját.
riporter gén- olyan gén, amelynek termékét egyszerű és érzékeny módszerekkel határozzák meg, és amelynek aktivitása a vizsgált sejtekben általában hiányzik. Génmanipulált konstrukciókban használják a vektor jelenlétének megerősítésére.
génfokozó(enhancer) - egy rövid DNS-szegmens, amely befolyásolja bizonyos gének megnyilvánulásának (kifejeződésének) szintjét, növelve az iniciáció és a transzkripció gyakoriságát.
heterozigóta- egy sejt (vagy szervezet), amely két különböző allélt tartalmaz a homológ kromoszómák egy adott lokuszán.
Heterozigótaság- különböző allélok jelenléte egy diploid sejtben.
heterozigóta szervezet olyan organizmus, amelynek kettő van különféle formák egy adott gén (különböző allélok) homológ kromoszómákon.
Heterokromatin- a kromoszóma olyan régiója (néha egy teljes kromoszóma), amely a transzkripció hiánya miatt interfázisban sűrű kompakt szerkezettel rendelkezik.
in situ hibridizáció- az egyszálú RNS vagy DNS radioaktív izotópjaival vagy immunfluoreszcens vegyületeivel jelölt sejtek denaturált DNS-e közötti hibridizáció üveglemezen.
DNS hibridizáció- kettős szálú DNS vagy DNS:RNS duplexek kialakulása a kísérlet során komplementer nukleotidok kölcsönhatása következtében.
Szomatikus sejtek hibridizációja- nem ivaros sejtek fúziója, szomatikus hibridek előállításának módszere (lásd).
hibrid fehérje (polipeptid) - lásd Fúziós fehérje (polipeptid).
hibridómák- hibrid limfoid sejtek, amelyeket egy tumor mielóma sejtnek immunizált állat vagy személy normál limfoid sejtjeivel való fúziójával nyernek.
Glikoziláció- szénhidrát-maradék rögzítése a fehérjéhez.
Holland örökség- Y-kapcsolt öröklődés.
Homozigóta- egy sejt (vagy szervezet), amely két azonos allélt tartalmaz a homológ kromoszómák egy adott lokuszában.
Homozigozitás- azonos allélok jelenléte diploid sejtben.
Homozigóta szervezet Olyan organizmus, amelynek homológ kromoszómáiban egy adott gén két azonos másolata található.
homológ kromoszómák Kromoszómák, amelyek ugyanazzal a génkészlettel rendelkeznek, mint az őket alkotó.
kuplung csoport Minden gén ugyanazon a kromoszómán található.
Genetikai ujjlenyomat- a tandem DNS ismétlődések számának és hosszának eltéréseinek kimutatása.
törlés- a kromoszómamutáció egy fajtája, amelyben a kromoszóma egy része elvész; a génmutáció olyan típusa, amelyben a DNS-molekula egy része hiányzik.
Denaturáció- a molekula térszerkezetének megsértése az intra- vagy intermolekuláris nem kovalens kötések megszakadása következtében.
Dihibrid kereszt- olyan organizmusok keresztezése, amelyek két alternatív karakterpárban különböznek egymástól, például a virág színe (fehér vagy színes) és a mag alakja (sima vagy ráncos).
DNS polimeráz- egy enzim, amely a DNS templát szintézisét vezeti.
hazai gének(Housekeeping gén) - ezek olyan gének, amelyek relatív állandósággal íródnak át, és normalizálóként (standard) használatosak a PCR-ben (polimeráz láncreakció), mivel feltételezzük, hogy expressziójukat nem befolyásolják a kísérleti körülmények.
dominancia- csak egy allél domináns megnyilvánulása egy tulajdonság kialakulásában egy heterozigóta sejtben.
Uralkodó- heterozigótákban megnyilvánuló tulajdonság vagy megfelelő allél.
Génsodródás- a génfrekvenciák változása számos generációban a mitózis, a megtermékenyítés és a szaporodás véletlenszerű eseményei miatt.
megkettőzés- a kromoszómamutáció olyan típusa, amelyben a kromoszóma bármely része megkétszereződik; egyfajta génmutáció, amelyben egy DNS-darab megkettőződik.
Genetikai próba- ismert szerkezetű vagy funkciójú DNS vagy RNS rövid darabja, amely valamilyen radioaktív vagy fluoreszcens vegyülettel van megjelölve.
Változékonyság- a tulajdonságok változatossága (diverzitása) e faj képviselői között.
Immunitás- a szervezet fertőző ágensekkel, például vírusokkal és mikrobákkal szembeni harcának mechanizmusa.
Immuntoxin- komplex egy antitest és bármely fehérjeméreg katalitikus alegysége között (diftériatoxin, ricin, abrin stb.).
Immunfluoreszcens szondák- lásd DNS-próbák, RNS-próbák.
Induktor- olyan faktor (anyag, fény, hő), amely inaktív állapotban lévő gének transzkripcióját okozza.
Profág indukció- a fág vegetatív fejlődésének beindítása a lizogén sejtekben.
Integrase- olyan enzim, amely meghatározott helyen keresztül genetikai elemet visz be a genomba.
integronok- olyan genetikai elemek, amelyek egy integráz gént, egy specifikus helyet és mellette egy promotert tartalmaznak, ami lehetővé teszi számukra, hogy mobil génkazettákat integráljanak magukba és kifejezzék a bennük lévő promóter nélküli géneket.
Interferonok- a gerincesek sejtjei által szintetizált fehérjék vírusfertőzésre válaszul, és elnyomják fejlődésüket.
Intron- egy gén nem kódoló régiója, amelyet átírnak, majd eltávolítanak az mRNS-prekurzorból a splicing szerkesztés során.
Intronizált gén- intronokat tartalmazó gén.
Iterons- a DNS-ben lévő nukleotid-maradékok ismétlődő szekvenciái.
Callus- a növény károsodásakor keletkező differenciálatlan sejtek tömege. Mesterséges táptalajon történő tenyésztésük során egyedi sejtekből képződhet.
kapszid a vírus fehérjeköpenyét.
Kifejezési kazetta- DNS-fragmentum, amely tartalmazza a bejuttatott gén kifejeződéséhez szükséges összes genetikai elemet.
cDNS- egyszálú DNS szintetizálva in vivo az RNS templáton reverz transzkriptáz segítségével.
Klón- genetikailag azonos sejtek csoportja, amelyek aszexuálisan egy közös őstől származnak.
KlónozásDNS- rekombináns DNS-molekulák előállításának folyamata idegen DNS-nek vektor-DNS- vagy RNS-molekulába ágyazásával, és ennek a konstrukciónak a fág-, bakteriális vagy eukarióta gazdasejtekbe történő bejuttatásával.
Klónozássejteket- szétválasztásuk tápközegben történő szitálás és izolált sejtből származó utódot tartalmazó telepek kinyerésével.
kodon- a DNS-ben vagy RNS-ben három egymást követő nukleotid-maradék, amely egy specifikus aminosavat kódol, vagy a transzláció végét jelzi.
Feldarabolás- a folyamat (termék) korlátozása a sejt egy bizonyos területére.
Kompetencia- a sejtek átalakulási képessége.
komplementaritás(a genetikában) - a nitrogénbázisok azon tulajdonsága, hogy a nukleinsavláncok kölcsönhatása során hidrogénkötések segítségével adenin-timin (vagy uracil) és guanin-citozin páros komplexeit képezzék.
konkatemer DNS- lineáris DNS, amelyben néhány elem (például a fág genom) többször megismétlődik.
contig- a szekvenálás során több, egymás után összefüggő DNS-szakasz csoportja.
konjugált- több kovalens kötésű molekulából álló komplexum.
Konjugáció- baktériumokban a genetikai információcsere módszere, amelynek során a sejtek közötti fizikai érintkezés következtében sejtes, plazmid vagy transzpozon DNS kerül át donor sejtből a recipiens sejtbe.
Cosmid- a λ fág cos-site DNS-ét tartalmazó vektor.
Átkelés- a homológ kromoszómák metszeteinek kicserélődésének jelensége a konjugáció során a meiózis során.
lektinek- a szénhidrátokat megkötő fehérjék.
Ligaz- egy enzim, amely foszfodiészter kötést hoz létre két polinukleotid között.
ligandum- egy meghatározott szerkezet által felismert molekula, például egy sejtreceptor.
Vezető sorozat- Szekretált fehérjék N-terminális szekvenciája, amely biztosítja a membránon keresztüli transzportjukat és egyúttal le is hasad.
Lysis- a sejt szétesése, amelyet héjának megsemmisülése okoz.
Lizogén- az a jelenség, hogy egy fágot prófág formájában hordoznak bakteriális sejtek (lásd: profág).
sejtvonal- genetikailag homogén állati vagy növényi sejtek, amelyek termeszthetők in vitro határozatlan ideig.
Linker- DNS-fragmensek összekapcsolására használt rövid szintetikus oligonukleotid in vitro; általában tartalmaz egy felismerési helyet egy specifikus restrikciós enzim számára.
ragadós végek- A DNS-molekulák végein elhelyezkedő, komplementer egyszálú DNS-szakaszok.
Liposzómák- mesterséges membránnal körülvett folyadékcseppek; mesterséges lipid vezikulák (lásd vezikulák).
A fág litikus fejlődése- a fág életciklusának fázisa, amely a sejt fertőzésével kezdődik és annak lízisével ér véget.
Locus- DNS-szakasz (kromoszómák), ahol egy bizonyos genetikai determináns található.
marker gén- egy gén a rekombináns DNS-ben, amely szelektív tulajdonságot kódol.
anyai hatásgének- a tojásban megjelenő gének, amelyek meghatározzák az utódok fenotípusát, függetlenül a hím genotípusától.
Interspecifikus hibridek- különböző fajokhoz tartozó sejtek fúziójából nyert hibridek.
Anyagcsere- a sejt létezését és szaporodását biztosító enzimfolyamatok összessége.
Metabolit- élő sejt kémiai reakciói során keletkező anyag.
Metiláz- enzimek, amelyek metilcsoportot kapcsolnak a DNS bizonyos nitrogénbázisaihoz.
mikroműhold- mikroszatellit lókusz (STR - az angol Short Tandem Repeats szóból): specifikus genomi lokalizációjú DNS-szakasz, amely rövid tandem ismétlődéseket tartalmaz.
minicellák- kromoszómális DNS-t nem tartalmazó sejtek. A biopolimer módosítása a szerkezetének megváltoztatása.
A genom mobil elemei- Olyan DNS-szekvenciák, amelyek az élő szervezetek genomjában mozoghatnak.
monohibrid kereszt- keresztezési formák, amelyek egy pár alternatív tulajdonságban különböznek egymástól.
Monoklonális antitestek- hibridómák által szintetizált, egy adott antigénre specifikus antitestek (lásd hibridómák).
Morfogenezis- a szervezet fejlesztését szolgáló genetikai program végrehajtása.
Mutagenezis- a mutációk indukciós folyamata.
Mutagének- fizikai, kémiai vagy biológiai ágensek, amelyek növelik a mutációk gyakoriságát.
Mutáció- a genetikai anyag megváltozása, amely gyakran a szervezet tulajdonságainak megváltozásához vezet.
Mouton- a mutáció elemi egysége, azaz a genetika legkisebb parcellája. anyag, a változás to-rogo azt a mutációt képviseli, amely fenotípusosan fogott és diszfunkcióhoz vezet. gén.
Öröklés- a genetikai információ átvitelének folyamata az organizmusok egyik generációjáról a másikra.
Átöröklés- az élőlények azon tulajdonsága, hogy anyagi és funkcionális folytonosságot biztosítsanak a generációk között, valamint megismételjenek egy bizonyos típusú egyedfejlődést.
örökölhetőség- a fenotípusos variabilitás aránya egy populációban a genetikai változatosság miatt (egy bizonyos minőségi vagy mennyiségi tulajdonsághoz viszonyítva).
Nick- egyszálú szakadás a DNS-duplexben 3'OH- és 5'p-végek képződésével; DNS-ligáz eliminálja (lásd DNS-ligáz).
Nitrogenáz egy olyan enzim, amely megköti a légköri nitrogént.
Nukleázok - gyakori név nukleinsavakat lebontó enzimek.
Reverz transzkriptáz- egy enzim, amely katalizálja a DNS-szintézis reakcióját egy RNS-templáton.
Oligonukleotid- DNS-lánc, amely több (2-20) nukleotid-maradékból áll.
Onkogének- olyan gének, amelyek termékei képesek az eukarióta sejteket úgy átalakítani, hogy elnyerjék a tumorsejtek tulajdonságait.
oncornavírus- RNS-tartalmú vírus, amely a normál sejtek rákos sejtekké való degenerálódását okozza; reverz transzkriptázt tartalmaz.
Operátor- a gén szabályozó régiója (operon), amely specifikusan megköti a represszort (lásd represszor), ezáltal megakadályozza a transzkripció megindulását.
Operon- együtt átíródó gének halmaza, amelyek általában a kapcsolódó biokémiai funkciókat szabályozzák.
Plazmid- körkörös vagy lineáris DNS-molekula, amely autonóm módon replikálódik a sejtkromoszómából.
Polilinker- egy szintetikus oligonukleotid, amely több restrikciós enzim felismerési helyeit tartalmazza (lásd restrikciós enzim).
Polimerázok- a nukleinsavak mátrixszintézisét vezető enzimek.
polipeptid- fehérje, peptidkötésekkel összekapcsolt aminosav-maradékokból álló polimer.
alapozó- egy rövid oligo- vagy polinukleotid szekvencia szabad 3'OH-csoporttal, amely komplementeren kapcsolódik egyszálú DNS-hez vagy RNS-hez; 3'-végéről a DNS polimeráz elkezd polidezoxiribonukleotid láncot építeni.
prokarióták sejtmaggal nem rendelkező organizmusok.
promóter- a gén szabályozó régiója (operon), amelyhez az RNS-polimeráz kötődik, hogy megindítsa a transzkripciót.
Proto-onkogének- normál kromoszómális gének, amelyek mutációi a sejt rosszindulatú degenerációjához vezethetnek.
Protoplaszt Növényi vagy mikrobiális sejt, amelynek nincs sejtfala.
Prophage- a fág intracelluláris állapota olyan körülmények között, amikor lítikus funkciói elnyomódnak.
Feldolgozás- a módosítás speciális esete (lásd a módosítást), amikor a biopolimerben a láncszemek száma csökken.
jel- szerkezeti jellemzők a szervezet bármely szintjén
Polimerizmus- nem allél több gén kölcsönhatása, amely egyirányúan befolyásolja ugyanazon tulajdonság kialakulását; egy tulajdonság megnyilvánulásának mértéke a gének számától függ. A polimer géneket ugyanazokkal a betűkkel jelöljük, és ugyanazon lókusz alléljainak ugyanaz az alsó indexe.
Pleiotrópia- a többszörös génhatás jelensége. Egy gén azon képességében fejeződik ki, hogy több fenotípusos tulajdonságot befolyásol.
Regulon- a genomban szétszórt, de egy közös szabályozó fehérje alá tartozó génrendszer.
A génexpresszió szabályozása- a sejtek szerkezetének és működésének ellenőrzése, valamint a sejtdifferenciálódás, morfogenezis és adaptáció alapjai.
Rekombináns DNS-molekula(a génsebészetben) - egy vektor és egy idegen DNS-fragmens kovalens kombinációjának eredményeként nyerik.
Rekombináns plazmid- idegen DNS fragmentum(oka)t tartalmazó plazmid.
rekombináns fehérje- rekombináns DNS-molekulából történő expresszió eredményeként kapott fehérje, amelyet gyakran Escherichia coli-ból nyernek.
Rekombinációin vitro - tevékenységek in vitro rekombináns DNS-molekulák létrehozásához vezet.
Rekombináció homológ- genetikai anyag cseréje két homológ DNS-molekula között.
Helyspecifikus rekombináció- asszociáció két DNS-molekula vagy egy molekula szakaszának megszakításával és egyesítésével, amely bizonyos helyeken történik.
Recon- a genetika elemi egysége rekombináció, azaz min. genetikai terület anyag, amelyen belül a rekombináció lehetséges.
Renaturáció- a molekulák eredeti térszerkezetének helyreállítása.
DNS javítás- a DNS-molekula károsodásának helyreállítása, eredeti szerkezetének helyreállítása.
replikátor a DNS azon régiója, amely a replikáció megindításáért felelős.
replikáció- a nukleinsavmolekulák megkettőzésének folyamata.
Replikon- replikátor irányítása alatt álló DNS-molekula vagy annak szakasza.
Elnyomás- a génaktivitás elnyomása, leggyakrabban transzkripciójuk blokkolásával.
Represszor- fehérje vagy antiszensz RNS, amely elnyomja a gének aktivitását.
Restriktázok- bakteriális helyspecifikus endonukleázok csoportja, amelyek felismerik a DNS bizonyos szakaszait négy vagy több nukleotidpár hosszúságban, és elhasítják a nukleotidláncot a felismerési helyen belül vagy kívül, "ragadós" vagy "tompa" végeket képezve.
Korlátozások- Restrikciós enzim által végzett hidrolízise után keletkezett DNS-fragmensek.
Korlátozási kártya- egy DNS-molekula diagramja, amely bemutatja a különböző restrikciós enzimekkel történő vágás helyeit.
Restrikciós elemzés- DNS hasítási helyek kialakítása restrikciós enzimekkel.
Retrovírusok- RNS-tartalmú állati vírusok, amelyek reverz transzkriptázt kódolnak és kromoszómális lokalizációjú provírust képeznek.
recesszív- az allél nem vesz részt egy tulajdonság kialakulásában heterozigóta sejtben.
Ribonukleázok (RNáz) – az RNS-t emésztő enzimek.
Szűrés- a rekombináns DNS-molekulákat tartalmazó telepek keresése szűrősejtekben vagy fágokban.
Weboldal- DNS-molekula, fehérje, stb.
Szekvenálás- nukleinsavak vagy fehérjék (polipeptidek) molekuláiban a linkek sorrendjének megállapítása.
Szelektív média- Tápközeg, amelyen csak bizonyos tulajdonságokkal rendelkező sejtek növekedhetnek.
Válaszfal- az osztódási ciklus végén a baktériumsejt közepén kialakuló és azt két leánysejtre osztó szerkezet.
Fúziós fehérje (polipeptid) egy fehérje, amely két különböző polipeptid fúziójával képződik.
Szomatikus hibridek nem nemi sejtek fúziójának terméke.
szomatikus sejtek- többsejtű szervezetek szöveteinek sejtjei, amelyek nem kapcsolódnak a nemhez.
távtartó- DNS-ben vagy RNS-ben - gének közötti nukleotidok nem kódoló szekvenciája; a fehérjékben egy aminosavszekvencia, amely a szomszédos globuláris doméneket kapcsolja össze.
Illesztés- érett mRNS vagy funkcionális fehérje képződésének folyamata a molekulák belső részeinek - az RNS intronoknak vagy inteineknek a fehérjékből történő eltávolításával.
Szuperproducer- egy mikrobiális törzs, amely egy bizonyos termék magas koncentrációban történő szintézisére irányul.
transzdukció- DNS-fragmensek átvitele bakteriofág segítségével.
Átírás- RNS-szintézis DNS-templáton; RNS polimeráz végzi.
átirat- egy transzkripciós termék, azaz egy adott DNS-helyen, mint egy templáton szintetizált RNS, amely komplementer annak egyik szálával.
Transzkriptáz fordított- egy enzim, amely az RNS-ből templátként a vele komplementer egyszálú DNS-t szintetizálja.
Adás- a fehérjék polipeptidláncának szintézise riboszómákban.
transzpozon- olyan genetikai elem, amely replikon részeként replikálódik, és önálló mozgásra (transzpozícióra) és a kromoszómális vagy extrakromoszómális DNS különböző részeibe történő integrálódásra képes.
Transzfekció- sejtek transzformációja izolált DNS felhasználásával.
átalakítás- az abszorbeált DNS okozta változás a sejt örökletes tulajdonságaiban.
átalakítás(molekuláris genetikában) - a genetikai információ átadása izolált DNS-en keresztül.
átalakítás(onkotranszformáció) - a sejtek részleges vagy teljes dedifferenciálódása, amelyet a sejtnövekedés diszregulációja okoz.
mérsékelt égövi fág- egy bakteriofág, amely képes egy sejt lizogenizálására, és a bakteriális kromoszómán belül vagy plazmid állapotban prófég formájában van.
F tényező (termékenységi tényező, nemi tényező) - a sejtekben található konjugatív F-plazmid E. coli(lásd ragozás).
Fenotípus- a szervezet tulajdonságainak külső megnyilvánulása, genotípusától és környezeti tényezőktől függően.
Kimérák- laboratóriumi hibridek (rekombinánsok).
Kromatin- a dezoxiribonukleoprotein (DNP) fonalas komplex molekulái, amelyek hisztonokhoz kapcsolódó DNS-ből állnak.
Centromere- egy kromoszómán lévő lókusz, amely fizikailag szükséges a homológ kromoszómák leánysejtek közötti eloszlásához.
Shine-Dalgarno sorozat- a prokarióta mRNS egy szakasza, amely a riboszóma rászállásához és megfelelő transzlációjához szükséges. A 16S riboszomális RNS 3' végével komplementer nukleotidszekvenciát tartalmaz.
DNS-keverés- két vagy több homológ fehérje génfragmenseinek rekombinációja. Három lépésből álló folyamat, amely magában foglalja a szülői DNS-molekulák megsemmisítését és két amplifikációs kört (primerek nélkül és speciálisan kiválasztottakkal), hogy hosszúságú, de megváltozott összetételű (kevert szekvenciákkal) kiméra DNS-molekulákat kapjunk, jelentősen javítva. vagy az általuk kódolt fehérjék új tulajdonságait
Szűrd le- sejtek, baktériumok (vagy vírusok) sora, amely egyetlen sejtből (vagy vírusból) vezet.
 — Születésnapok gyerekeknek és felnőtteknek
— Születésnapok gyerekeknek és felnőtteknek Családi kávézó csokoládé lány kávézókban és éttermekben
Családi kávézó csokoládé lány kávézókban és éttermekben — Születésnapok gyerekeknek és felnőtteknek
— Születésnapok gyerekeknek és felnőtteknek