A kromoszómaelmélet alapjai. Morgan kromoszómaelmélete: meghatározás, alapok és jellemzők
Kromoszóma-elmélet kialakulása 1902-1903. W. Setton amerikai amerikai citológus és T. Bo-Veri német citológus és embriológus önállóan felfedezett párhuzamosságot a gének és kromoszómák viselkedésében a gamete képződése és megtermékenyítése során. Ezek a megfigyelések szolgáltak a feltételezéshez, hogy a gének a kromoszómákban helyezkednek el. A specifikus gének specifikus kromoszómákban való lokalizációját azonban csak 1910-ben szerezte meg T. Morgan amerikai genetikus, aki az ezt követő években (1911–1926) igazolta az öröklési kromoszóma elméletet. Ezen elmélet szerint az örökletes információ átvitele kromoszómákhoz kapcsolódik, ahol a gének lineárisan, bizonyos szekvenciában lokalizálódnak. Így az öröklés anyagi alapját a kromoszómák képviselik.
A kromoszóma-elmélet kialakulását megkönnyítette a nemi genetika vizsgálatával nyert adatok, amikor a különböző nemű szervezetekben a kromoszómák halmazának különbségeit állapították meg.
A homológ kromoszómák konjugálása során a meiosis I. prófázájában a crossover előfordul (3.10. Ábra). Ekkor a két kromoszóma részei metszhetnek és kicserélhetik részeiket. Ennek eredményeképpen kvalitatívan új kromoszómák keletkeznek, amelyek mind anyai, mind apai kromoszómák metszeteit tartalmazzák. egyének, az allélok új kombinációjával kapott ilyen gamétákból származó keresztmetszet vagy rekombináns.
Az ugyanabban a kromoszómában található két gén metszéspontjának gyakorisága (százalékos aránya) arányos a közöttük lévő távolsággal. A két gén közötti átkelés ritkábban fordul elő, annál közelebb vannak egymáshoz. Ahogy a gének távolsága növekszik, nő a valószínűség, hogy az átkelés két különböző homológ kromoszómán keresztül oszlik meg.
A gének közötti távolság jellemzi a markolat erősségét. Vannak olyan gének, amelyek nagy arányban vannak a fogással, és azok, ahol a tapadás szinte észrevehetetlen. Összekapcsolt örökséggel azonban a crossover maximális értéke nem haladja meg az 50% -ot. Ha magasabb, akkor egy szabad kombináció van az allélpárok között, amelyek nem különböznek a független öröklésektől.
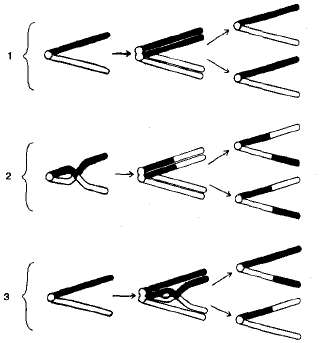
Ábra. 3.10. Crossover rendszer: I - az átkelés hiánya; 2 - átkelés két kromoszóma színpadán; 3 - áthalad a négy kromatid színpadán.
Tekintsük T. Morgan egyik első kísérletét a kapcsolt öröklés tanulmányozására. A gyümölcs legyek átlépésekor, két pár kapcsolt jelzésű párban - szürke, kezdeti szárnyakkal és fekete normál szárnyakkal - eltérő hibridek F 1 szürke volt a normál szárnyakkal (3.11. ábra).
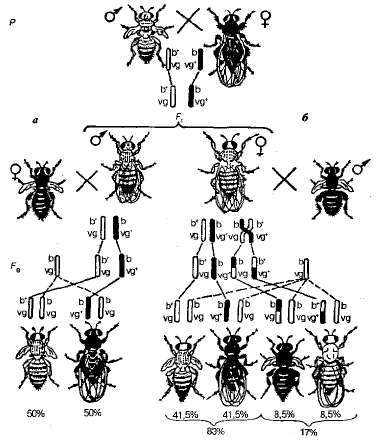
Ábra. 3.11. Drosophilában a párosított tulajdonságok öröksége: teljes tapadás (hímnemű átkelés nélküli digerozozigrten nélkül); b - összekapcsolás crossoverrel (diheterozigattos nő, amelyben az átkelés nincs elnyomva); b +, b - ~ szürke és fekete test színe; vg + , vg - - normál és kezdeti szárnyak.
Továbbá kétféle elemző keresztet hajtottunk végre. Az elsőben diheterozigóta hímeket vettünk. F 1és a recesszív allélekre homozigóta nőstényekkel, és a másodikban a diheterozigóta nőstényeket a férfiakkal recesszívák mindkét tulajdonságra (fekete test és kezdeti szárnyak). E kereszteződések eredményei eltérőek voltak (lásd 3.11. Ábra).
Az első esetben ennek a kísérletnek a szülői (P) szülői fenotípussal való leszármazottai, vagyis a szürke szárnyak kezdetleges szárnyakkal és fekete normál szárnyakkal repül 1: 1 arányban. Ezért ez a diheterozigóta csak kétféle ivarsejtet alkot. (b + vg és bvg +) négy helyett. E hasítás alapján feltételezhetjük, hogy a hímnek teljes génkötése van.
A második esetben -ban F 2 egy másik hasítást figyeltünk meg. A szülői kombinációk mellett újfajta jelek is megjelennek - fekete testtel és alapos szárnyakkal, valamint szürke testtel és normál szárnyakkal repülnek. A rekombináns leszármazottak száma azonban kicsi és 17%, a vezetők száma 83%. Az új karakterek kombinációjával rendelkező kis legyek megjelenésének oka áthalad, ami a gének alléljainak új rekombináns kombinációjához vezet. b és vg homológ kromoszómákban. Ezek a cserék 17% -os valószínűséggel fordulnak elő, és végső soron két valószínűséggel azonos rekombináns osztályt adnak - mindegyik 8,5%.
Az átkelés biológiai jelentősége rendkívül magas, mivel a genetikai rekombináció lehetővé teszi új, korábban nem létező gének kombinációinak létrehozását, és ezáltal az örökletes variabilitást, amely számos lehetőséget kínál a szervezet különböző környezeti feltételekhez való alkalmazkodására. Egy személy kifejezetten hibridizálást végez annak érdekében, hogy a tenyésztési munkákhoz szükséges lehetőségek kombinációját megkapja.
A genetikai térkép fogalma. T. Morgan és munkatársai K. Bridges, A. Stertevanti G. Moler kísérletileg kimutatták, hogy a kapcsolás és az átkelés jelenségeinek ismerete nemcsak a gének összekapcsoló csoportjának létrehozását teszi lehetővé, hanem a kromoszómák genetikai térképeinek megalkotását is, amelyeken a kromoszómában és a rokonban lévő gének sorrendje a távolság közöttük.
Genetikai kromoszóma térkép az azonos kapcsolati csoportban lévő gének kölcsönös elrendezésének rendszerét nevezzük. Az ilyen térképeket minden homológ kromoszóma párra állítjuk össze.
Az ilyen leképezés lehetősége az egyes gének közötti átkelés százalékos állandóságán alapul. A kromoszómák genetikai térképeit számos organizmusra állítják össze: rovarok (Drosophila, szúnyog, csótány stb.), Gombák (élesztő, aspergillus), baktériumok és vírusok esetében.
A genetikai térkép jelenléte azt jelzi, hogy egy vagy másik típusú szervezet nagyfokú tanulmányokat végez, és nagy tudományos érdeklődéssel bír. Egy ilyen szervezet kiváló kísérleti munkát jelent, amely nem csak tudományos, hanem gyakorlati jelentőséggel bír. Különösen a genetikai térképek ismerete lehetővé teszi, hogy terveket dolgozzon ki a jelek bizonyos kombinációival rendelkező szervezetek megszerzésére, amelyet ma már széles körben használnak a tenyésztési gyakorlatban. Így a farmakológia és a mezőgazdaság számára szükséges fehérjék, hormonok és más komplex szerves anyagok szintetizálására alkalmas mikroorganizmus törzsek létrehozása csak a géntechnológiai módszerek alapján lehetséges, amelyek viszont a megfelelő mikroorganizmusok genetikai térképének ismeretén alapulnak.
Az emberi genetikai térképek hasznosak lehetnek az egészségügyben és az orvostudományban is. A gén lokalizációjának ismerete egy adott kromoszómában számos súlyos örökletes betegség diagnosztizálására szolgál. Már létezik génterápiás lehetőség, azaz a gének szerkezetének vagy funkciójának javítása.
Az élő szervezetek különböző fajainak genetikai térképeinek összehasonlítása hozzájárul az evolúciós folyamat megértéséhez.
Az öröklési kromoszóma elmélet főbb rendelkezései. A kapcsolt öröklés jelenségeinek elemzése, átkelés, genetikai és citológiai térképek összehasonlítása lehetővé teszi, hogy megfogalmazzuk az öröklési kromoszóma elmélet főbb rendelkezéseit:
- A gének a kromoszómákban lokalizálódnak. Ugyanakkor a különböző kromoszómák különböző számú gént tartalmaznak. Ezenkívül az egyes nem homológ kromoszómák gének halmaza egyedülálló.
- Az allélgének ugyanazon lokuszokat foglalják el a homológ kromoszómákban.
- A gének lineáris sorrendben helyezkednek el a kromoszómában.
- Ugyanazon kromoszóma gének egy kötőcsoportot képeznek, aminek következtében bizonyos jelek kapcsolt öröksége keletkezik. Ebben az esetben a tapadás ereje fordítottan kapcsolódik a gének közötti távolsághoz.
- Mindegyik fajra jellemző a kromoszómák meghatározott csoportja - a kariotípus.
Forrás : NA Lemeza L.V. Kamluk N.D. Lisov "Biológiai kézikönyv az egyetemekbe való belépéshez"
A Morgan munkája megalapozta az öröklési kromoszóma-elmélet alapjait, kimutatta, hogy egyes gének szabad kombinatorikájának korlátai a gének egy kromoszómában való elhelyezkedésének és fizikai kötődésének köszönhetők.
Morgan megállapította, hogy az ugyanazon kromoszómán található gének kapcsolata nem abszolút. A meiosis során az egyik páros kromoszómák egymás között homológ helyeket cserélhetnek egy átkeléses eljárással. Minél távolabb vannak egymástól a kromoszómában levő gének, annál gyakrabban kerülnek átadásra az átkelés során. E jelenség alapján a gén-kohézió mértékét javasolták - az átkelés százalékos arányát -, és meghatározták a különböző Drosophila fajok kromoszómáinak első genetikai térképeit.
A genetikai elemzés tárgyaként Drosophila gyümölcseit választották, és Morgan különböző vonások örökségét tanulmányozta benne.
Homozigóta nőstény átkelése szürke testtel és hosszú szárnyakkal (domináns), homozigóta, fekete szárnyú rövid szárnyú hím, F1-ben - egyenletesség (szürke test, hosszú szárnyak)
Kiderült, hogy az eredmények a hibrid nemétől függően eltérőek lesznek.
Ha a hím hibrid volt, akkor az utódok két fenotípusos osztályt termeltek, amelyek teljes mértékben megismételték a szülők jellemzőit.
Ha a nő hibrid volt, akkor 4 leszármazott fenotípusos osztály létezett egyenlőtlen arányban. Az utódok többsége (83%) szülői tulajdonságokkal rendelkező, utóbbiak, kevesebb (17%) - új tulajdonságokkal rendelkező egyének.
Morgan arra a következtetésre jutott, hogy a tengelykapcsoló hiányos lehet, ha a tengelykapcsoló-csoportot áthidalással megszakítják.
A leszármazottak szokatlan százalékát azzal magyarázza, hogy az átkelés nem mindig történik meg, az átkelés gyakorisága függ a gének közötti távolságtól - minél hosszabb a távolság, annál kisebb a gén közötti összekapcsoló erő, annál gyakrabban az átkelés.
A nem keresztezett kromoszómákkal rendelkező gametákat nem crossovernek nevezik.
Ha a gametákban a kereszteződésen átesett kromoszómák átkelnek.
6. Az öröklési kromoszóma elmélet főbb rendelkezései
1. A gének a kromoszómákban lineárisan helyezkednek el bizonyos területeken - loci. Az allélgének azonos homológ kromoszómák helyét foglalják el.
2. Az ugyanabban a kromoszómában található gének összekötő csoportot alkotnak, és együtt öröklődnek vagy összekapcsolódnak. A kapcsolócsoportok száma = a haploid készletben lévő kromoszómák száma.
3. A homológ kromoszómák közötti áthidalás lehetséges, megszakítva a kapcsolást
4. az átkelés folyamata közvetlenül arányos a gének közötti távolsággal.
1% -os átkelés = 1 Celsius
7. A citoplazmatikus öröklés fogalma
Egy bizonyos mennyiségű örökletes anyag jelenléte a citoplazmában a mitokondriumok és a plasztidok körkörös DNS-molekuláinak, valamint más, nukleáris genetikai elemeknek a formájában indokolja, hogy kifejezetten a fenotípus kialakulásában való részvételükre összpontosítson az egyéni fejlődés folyamatában.
A citoplazmatikus gének nem tartoznak a Mendeli öröklési törvények hatálya alá, amelyeket a kromoszómák viselkedése határoz meg a mitózisban, a meiózisban és a megtermékenyítésben. Annak a ténynek köszönhetően, hogy a megtermékenyítés eredményeképpen létrejött organizmus citoplazmatikus struktúrákat fogad el főként egy tojássejtrel, a tulajdonságok citoplazmatikus öröksége az anyai vonalon keresztül történik. Ezt a fajta örökséget először K. Correns 1908-ban írta le egyes növényekben a tarka levelek jellegzetességei tekintetében.
És megtermékenyítés. Ezek a megfigyelések szolgáltak a feltételezéshez, hogy a gének a kromoszómákban helyezkednek el. A specifikus gének lokalizációját kísérleti bizonyítékként csak specifikus kromoszómákban találtuk meg, csak a T. Morgan amerikai genetikus városban, aki a következő években (-) igazolta az öröklési kromoszóma elméletet. Ezen elmélet szerint az örökletes információ átvitele kromoszómákhoz kapcsolódik, ahol a gének lineárisan, bizonyos szekvenciában lokalizálódnak. Így az öröklés anyagi alapját a kromoszómák képviselik.
A kromoszóma-elmélet kialakulását megkönnyítette a nemi genetika vizsgálatával nyert adatok, amikor a különböző nemű szervezetekben a kromoszómák halmazának különbségeit állapították meg.
A padló genetikája
Hasonló módszer a nem (XY-típus) meghatározására minden emlősnek, beleértve az embereket is, akiknek sejtjei 44 autoszómát és két X kromoszómát tartalmaznak nőkben vagy XY kromoszómákat férfiaknál.
Így XY-típusú nemi meghatározás, vagy a gyümölcsfajta és az ember típusa, - a leggyakoribb módja a nemek meghatározásánakA legtöbb gerincesre és néhány gerinctelenre jellemző. Az X0-típus a legtöbb ortopteránban, bedugóban, bogárban, pókban található, ahol az Y-kromoszóma egyáltalán nem létezik, így a hím X0-genotípusa és a XX.
Minden madárban, a legtöbb lepkékben és néhány hüllőkben, a férfiak homogetikusak, a nők heterogémiásak (XY vagy XO típus). Ezeknek a fajoknak a nemi kromoszómáit Z és W betűkkel jelöljük, hogy ezt a nemi meghatározási módszert elkülönítsük; a férfiak kromoszómáit a ZZ szimbólum, a nőstények pedig a ZW vagy Z0 szimbólummal jelölik.
Bizonyíték arra, hogy a nemi kromoszómák Drosophilában történő nem-kromoszómáinak a vizsgálatát a nemi kromoszómák meghatározzák. Ha mindkét nemi kromoszóma bejut az egyik gametébe, és nem egybe a másikba, akkor az ilyen gamétáknak a normálokkal történő egyesítésekor a XXX, HO, XXY stb. és a HHU-val - nőkkel (az emberekben - az ellenkezőjével). A XXX csoporttal rendelkező egyének hipertrófiai jelei a nősténynek (szuper-nő). (Az összes kromoszóma-rendellenességgel rendelkező egyének Drosophilában sterilek). Később bebizonyosodott, hogy Drosophilában a nemet az X kromoszómák száma és az autoszómák száma között határozzák meg.
A nemhez kötődő tulajdonságok öröksége
Abban az esetben, ha egy adott tulajdonság képződését szabályozó gének autoszómákban vannak lokalizálva, az öröklés függetlenül attól függetlenül történik, hogy melyik szülő (anya vagy apa) a vizsgált tulajdonság hordozója. Ha a gének a nemi kromoszómákban vannak, a karakterek örökségének jellege drámaian változik. Például Drosophilában az X-kromoszómán lokalizált gének általában nem rendelkeznek allélekkel az Y-kromoszómán. Emiatt a heterogetikus szex X kromoszómájában a recesszív gének szinte mindig jelennek meg, mivel az egyedülálló.
A nemi kromoszómákban lokalizálódó jeleket szex-kapcsolt karaktereknek nevezik. A szexuális öröklés jelenségét T. Morgan fedezte fel Drosophilában.
Emberekben az X és Y kromoszómák homológ (pszeudoautoszomális) régióval rendelkeznek, ahol gének találhatók, amelyek öröksége nem különbözik az autoszomális gének örökségétől.
A homológ régiókon kívül az X és Y kromoszómák nemhomológiai régiókkal rendelkeznek. Az Y kromoszóma nem-homológ régiója a férfi nemet meghatározó gének mellett az emberi lábujjak és szőrös fülek közötti membránok génjeit tartalmazza. Az Y kromoszóma nonhomológ régiójához kapcsolódó patológiás jeleket minden fia átadja, mivel az apától az Y kromoszómát kapják.
Az X kromoszóma nem homológ régiója számos olyan gént tartalmaz, amelyek fontosak a szervezetek létfontosságú aktivitása szempontjából. Mivel az X kromoszóma heterogetett szex (XY) az egyedüli, az X kromoszóma nem homológ részének génjei által meghatározott jelek akkor is megjelennek, ha recesszívek. Ezt a gének állapotát hemizigótanak nevezik. Példa erre az X-kapcsolattal recesszív tünetek emberekben a hemofília, a Duchenne izomduzzanat, a látóideg atrófia, a színvakság stb
A hemofília olyan örökletes betegség, amelyben a vér elveszíti a vérrögképességét. A seb, még egy karcolás vagy zúzódás is, bőséges külső vagy belső vérzést okozhat, ami gyakran véget ér a halálnak. Ez a betegség ritka kivételekkel csak férfiaknál található. Azt tapasztaltuk, hogy mind a leggyakoribb hemofília (A hemofília, mind a hemofília B) formái miatt recesszív génekaz X kromoszómán található. Ezeknek a géneknek heterozigóta nők (hordozók) normális vagy enyhén csökkent véralvadást mutatnak.
A lányok hemofília fenotípusos megnyilvánulása akkor figyelhető meg, ha a lány anyja a hemofília gén hordozója, és az apa a hemofília. Hasonló öröklési mintázat jellemző a többi, recesszív szexuális tulajdonságra.
Kapcsolódó öröklés
A tulajdonságok független kombinációja (a Mendel harmadik törvénye) azzal a feltétellel történik, hogy az ilyen tulajdonságokat meghatározó gének különböző homológ kromoszómákban vannak. Következésképpen minden szervezetben a meiosisban önállóan kombinálódó gének száma korlátozódik a kromoszómák számával. A szervezetben azonban a gének száma jelentősen meghaladja a kromoszómák számát. Például több mint 500 gént vizsgáltak kukoricában a molekuláris biológia kora előtt, több mint ezer drosophila legyekben, és mintegy 2 ezer emberben, míg 10, 4 és 23 pár kromoszóma. W. Sutton számára a 20. század elején már egyértelmű volt, hogy a magasabb szervezetekben a gének száma több ezer volt. Ez azt sugallta, hogy sok kromoszóma lokalizálódik minden kromoszómán. Az ugyanabban a kromoszómában lokalizált gének összekötő csoportot alkotnak és együtt öröklődnek.
T. Morgan gének közös öröksége a kapcsolt öröklést hívta fel. A kapcsolócsoportok száma megfelel a kromoszómák haploid számának, mivel a kapcsolócsoport két homológ kromoszómából áll, amelyekben ugyanazok a gének találhatók. (A heterogetikus szexuális egyedeknél, például emlősök férfiaknál a tengelykapcsoló csoportok valójában még egy, mivel az X és Y kromoszómák különböző géneket tartalmaznak, és két különböző tengelykapcsolócsoportot tartalmaznak. férfiaknak - 24).
A kapcsolt gének öröklési módja eltér a homológ kromoszómák különböző párjaiban lokalizált gének örökségétől. Tehát, ha egy független kombinációban a diheterozigóta egyén négy fajta ivarsejtet (AB, Ab, aB és ab) egyenlő mennyiségben alkot, de láncolt örökléssel (az átkelés hiányában) ugyanaz a digerozigóta csak kétféle ivarsejtet alkot: (AB és ab) is egyenlő mennyiségben. Ez utóbbi megismétli a szülő kromoszómájában a gének kombinációját.
Megállapítást nyert azonban, hogy a szokásos (nem bokszos) gameták mellett más (cross-over) gameták vannak az Ab és aB gének új kombinációival, amelyek különböznek a szülő kromoszómáinak kombinációitól. Az ilyen ivarsejtek oka a homológ kromoszómák szakaszainak cseréje, vagy átkelés.
A homológ kromoszómák konjugálása során a meiosis I. prófázisa során a crossover előfordul. Ekkor a két kromoszóma részei metszhetnek és kicserélhetik részeiket. Ennek eredményeképpen kvalitatívan új kromoszómák keletkeznek, amelyek mind anyai, mind apai kromoszómák metszeteit tartalmazzák. Az ilyen gamétákból származó, az allélok új kombinációjával kapott egyedeket átkelésnek vagy rekombinánsnak nevezik.
Az ugyanabban a kromoszómában található két gén metszéspontjának gyakorisága (százalékos aránya) arányos a közöttük lévő távolsággal. A két gén közötti átkelés ritkábban fordul elő, annál közelebb vannak egymáshoz. Ahogy a gének távolsága növekszik, nő a valószínűség, hogy az átkelés két különböző homológ kromoszómán keresztül oszlik meg.
A gének közötti távolság jellemzi a markolat erősségét. Vannak olyan gének, amelyek nagy arányban vannak a fogással, és azok, ahol a tapadás szinte észrevehetetlen. Összekapcsolt örökség esetén azonban a maximális átviteli frekvencia nem haladja meg az 50% -ot. Ha magasabb, akkor egy szabad kombináció van az allélpárok között, amelyek nem különböznek a független öröklésektől.
Az átkelés biológiai jelentősége rendkívül magas, mivel a genetikai rekombináció lehetővé teszi új, korábban nem létező gének kombinációinak létrehozását, és ezáltal az örökletes variabilitást, amely számos lehetőséget kínál a szervezet különböző környezeti feltételekhez való alkalmazkodására. Egy személy kifejezetten hibridizálást végez annak érdekében, hogy a tenyésztési munkákhoz szükséges lehetőségek kombinációját megkapja.
A genetikai térkép fogalma
T. Morgan és munkatársai, K. Bridges, A. G. Sturtevant és G. J. Meller kísérletileg kimutatták, hogy a kötés és az átkelés jelenségeinek ismerete nemcsak a gének összekapcsoló csoportjának létrehozását teszi lehetővé, hanem a kromoszómák genetikai térképeinek megalkotását is, amelyeken a helyszín sorrendje szerepel. a kromoszómában lévő gének és a közöttük lévő relatív távolságok.
A kromoszómák genetikai térképét az azonos kapcsolati csoportban lévő gének kölcsönös elrendezésének sémájának nevezzük. Az ilyen térképeket minden homológ kromoszóma párra állítjuk össze.
Az ilyen leképezés lehetősége az egyes gének közötti átkelés százalékos állandóságán alapul. A kromoszómák genetikai térképeit számos organizmusra állítják össze: rovarok (Drosophila, szúnyog, csótány stb.), Gombák (élesztő, aspergillus), baktériumok és vírusok esetében.
A genetikai térkép jelenléte azt jelzi, hogy egy vagy másik típusú szervezet nagyfokú tanulmányokat végez, és nagy tudományos érdeklődéssel bír. Egy ilyen szervezet kiváló kísérleti munkát jelent, amely nem csak tudományos, hanem gyakorlati jelentőséggel bír. Különösen a genetikai térképek ismerete lehetővé teszi, hogy terveket dolgozzon ki a jelek bizonyos kombinációival rendelkező szervezetek megszerzésére, amelyet ma már széles körben használnak a tenyésztési gyakorlatban. Így a farmakológia és a mezőgazdaság számára szükséges fehérjék, hormonok és más komplex szerves anyagok szintetizálására alkalmas mikroorganizmus törzsek létrehozása csak a géntechnológiai módszerek alapján lehetséges, amelyek viszont a megfelelő mikroorganizmusok genetikai térképének ismeretén alapulnak.
Az emberi genetikai térképek hasznosak lehetnek az egészségügyben és az orvostudományban is. A gén lokalizációjának ismerete egy adott kromoszómában számos súlyos örökletes betegség diagnosztizálására szolgál. Már létezik lehetőség génterápiára, azaz a gének szerkezetének vagy funkciójának korrigálására.
Az öröklési kromoszóma elmélet főbb rendelkezései
A kapcsolt öröklés jelenségeinek elemzése, átkelés, genetikai és citológiai térképek összehasonlítása lehetővé teszi, hogy megfogalmazzuk az öröklési kromoszóma elmélet főbb rendelkezéseit:
- A gének a kromoszómákban lokalizálódnak. Ugyanakkor a különböző kromoszómák különböző számú gént tartalmaznak. Ezenkívül az egyes nem homológ kromoszómák gének halmaza egyedülálló.
- Az allélgének ugyanazon lokuszokat foglalják el a homológ kromoszómákban.
- A gének lineáris sorrendben helyezkednek el a kromoszómában.
- Ugyanazon kromoszóma génjei kötőcsoportot alkotnak, azaz öröklődnek, főként kapcsolt (együtt), aminek következtében a jelek egy része kapcsolódik. A tengelykapcsoló-csoportok száma megegyezik egy adott faj kromoszómáinak haploidszámával (a homogametikus szexben) vagy 1-nél (a heterogetikus nemben).
- Az áthidalás következtében a tapadás megszakad, amelynek frekvenciája közvetlenül arányos a kromoszóma gének közötti távolsággal (ezért a tapadás ereje fordítottan kapcsolódik a gének közötti távolsághoz).
- Mindegyik fajra jellemző a kromoszómák meghatározott csoportja - a kariotípus.
forrás
- N. A. Lemeza L. V. Kamlyuk N. D. Lisov „A biológiai kézikönyv az egyetemekbe való belépéshez”
jegyzetek
Wikimedia Alapítvány. 2010.
A kromoszómaelmélet (HT) alkotója Thomas Morgan tudós. A CT a sejtszintű öröklődés tanulmányozásának eredménye.
A kromoszóma-elmélet lényege:
Az öröklés anyagi hordozói kromoszómák.
Ennek fő bizonyítéka:
Citogenetikus párhuzamosság
Kromoszóma nemi meghatározás
A szexuális öröklés
Géncsatolás és átkelés
A chrosoma elmélet főbb rendelkezései:
Az örökletes lerakódások (gének) a kromoszómákon találhatók.
A gének lineárisan helyezkednek el a kromoszómában.
Mindegyik gén egy meghatározott régiót (lókuszt) foglal magában. Az allélgének hasonló lokuszokat foglalnak el a homológ kromoszómákban.
Az egyik kromoszómában lokalizált gének együtt öröklődnek, összekapcsolódnak (Morgan-törvény), és összekötő csoportot alkotnak. A tengelykapcsoló csoportok száma megegyezik a kromoszómák haploid számával (n).
A homológ kromoszómák cseréje vagy rekombináció között.
A gének közötti távolságot az átkelés százalékában mérjük - a morgánokat.
A keresztezési frekvencia fordítottan arányos a gének közötti távolsággal, és a gének közötti kohéziós erő fordítottan arányos a közöttük lévő távolsággal.
Citogenetikus párhuzamosság
A Morgan Sytton diplomája megjegyezte, hogy a Mendel szerint a gének viselkedése egybeesik a kromoszómák viselkedésével: (TABLE - citogenetikus párhuzamosság)
Mindegyik organizmus 2 örökletes betétet hordoz, a gamete csak egy-egy örökletes betétet tartalmaz. A zigótában és a testben további megtermékenyítés esetén 2 ismét örökletes betétek az egyes tulajdonságokhoz.
A kromoszómák ugyanúgy viselkednek, ami arra utal, hogy a gének a kromoszómákban vannak, és velük együtt örökölnek.
Kromoszóma nemi meghatározás
1917-ben Allen kimutatta, hogy a moha és a nőstény egyedek a kromoszómákban különböznek. A férfi test diploid szövetének sejtjeiben, a XiY nemi kromoszómákban, a női XiX-ben. Így a kromoszómák egy ilyen jelet nemekként definiálnak, és ezért az öröklődés anyagi hordozói lehetnek. Később a kromoszómális nemi meghatározást más szervezetek, köztük az emberek esetében is kimutatták. (TÁBLÁZAT)
A szexuális öröklés
Mivel a nemi kromoszómák férfi- és női szervezetekben eltérőek, az X- vagy Y-kromoszómákban található gének eltérő módon öröklik meg. Az ilyen jeleket hívják nemi kapcsolatok.
A nemhez kötődő tulajdonságok örökségének jellemzői
Nem figyelte meg 1 Mendel törvényét
A kölcsönös kereszteződések különböző eredményeket adnak.
Van egy kereszt-kereszt (vagy öröklési kereszt).
Először Morgan a Drosophilában fedezte fel a tulajdonsághoz kapcsolódó örökséget.
|
W + vörös szem |
(C) x w + x w + * x w y |
(C) x w x w * x w + y |
|||||
|
w - fehér szemek | |||||||
|
(CЖ) X W + X w - Vörös szem |
X w X W + - Vörös szemek |
||||||
|
(CM) X W + Y - Vörös szem |
X w Y– Fehér szemek |
||||||
|
Így a Morgan által azonosított mutáció öröksége - „fehér szem” - fehér volt, a fent felsorolt jellemzőkkel jellemezve: Az egységesség törvényét nem tartották be 2 kölcsönös kereszteződésben különböző utódok érkeztek A második kereszteződésben a fiúk az anya (fehér szemek), lányai - az apa jele (vörös szemek) jelzik. Az ilyen örökséget „ropogós kereszt örökségnek” nevezik. |
|||||||
(TÁBLÁZAT szexuális öröklés)
A szexuális öröklés az X-kromoszóma-génekhez allélikus Y-kromoszómában nem található gének hiánya, az Y-kromoszóma sokkal kisebb, mint az X-kromoszóma, 78 pedig most lokalizálódik benne (?) gének, míg kromoszómájukban több mint 1098.
Példák a nemi alapú öröklésre:
Hemophilia, Duchenne dystrophia, Duncan-szindróma, Alport-szindróma, stb.
Vannak olyan gének, amelyek ellenkezőleg, az Y-kromoszómában vannak, és nincsenek az X-kromoszómában, ezért csak férfias szervezetekben találhatók, soha nem a nőkben (holandrikus öröklés), és csak az apa fiaihoz kerülnek.
Géncsatolás és átkelés
A genetikában ismert egy olyan jelenség, mint a „gének vonzereje”: egyes nem-párhuzamos tulajdonságok nem öröklődtek egymástól függetlenül, mivel Mendel III. Törvénye, de együtt örökölt, nem adott új kombinációkat. Morgan ezt elmagyarázta azzal, hogy ezek a gének ugyanabban a kromoszómában vannak, így az egyik csoportban egymásba eső lánysejtekké válnak, mintha összekapcsolódnának. Ezt a jelenséget hívta - összefonódott örökség.
Morgan tengelykapcsoló törvény:
Az ugyanazon kromoszómában található gének együtt öröklődnek, összekapcsolódnak.
Az ugyanabban a kromoszómában található gének kötőcsoportot alkotnak. A tengelykapcsoló csoportok száma megegyezik az "n" - haploid kromoszómák számával.
Homozigóta legyek vonalakkal átléptek szürke testtel és hosszú szárnyakkal, és fekete testtel és rövid szárnyakkal repülnek. A test színei és a szárnyak hossza összekapcsolódik, azaz egy kromoszómában fekszenek.
|
A-szürke test egy fekete test B - normál szárnyak (hosszú) b - kezdeti szárnyak |
(CF) AABBxaabb (CM) |
|||||
|
Szürke hosszú szárnyas |
Fekete rövid szárnyú |
|||||
|
Jegyezze fel a kromoszóma-expressziót |
||||||
|
Szürke test Hosszú szárnyak |
Fekete test Rövid test |
|||||
|
|
|
|||||
|
Minden legyek egy szürke test és hosszú szárnyak. |
||||||
|
Ie ebben az esetben a hibridek I generációjának egységességét szabályozzák. Az F2-ben azonban a várt 9: 3: 3: 1-es osztás helyett 3 szürke hosszúszárnyú és 1-részű fekete rövid szárnyú arányt kaptunk; A jelek új kombinációi nem jelennek meg. Morgan azt javasolta, hogy a Deheterozygote F 2 - ( |
||||||
|
|
||||||
|
Szürke test Hosszú szárnyak |
Fekete test Rövid test |
|||||
|
|
|
|||||
|
F egy |
|
|
||||
|
Szürke test Hosszú szárnyak |
Fekete test Rövid szárnyak |
|||||
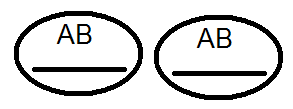
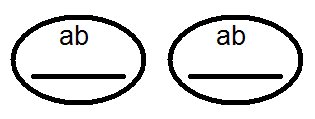
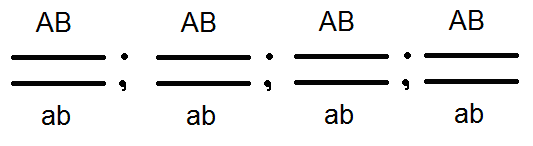
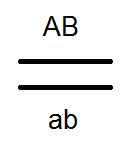
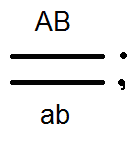 ) nem termel (nem) 4-et, de csak 2 fajtát -
) nem termel (nem) 4-et, de csak 2 fajtát -  és
és  . Az elvégzett keresztek elemzése ezt megerősítette:
. Az elvégzett keresztek elemzése ezt megerősítette: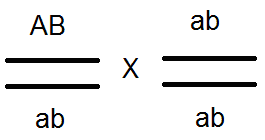
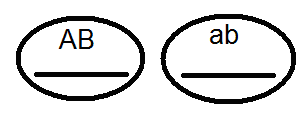
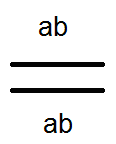
Ennek eredményeképpen az osztódás F2-ben történik, mint egy 3: 1-es monohibrid kereszteződésben.
|
|
||||
|
|
|
|||
|
|
|
|
|
|
|
Szürke test Hosszú szárnyak |
Szürke test Hosszú szárnyak |
Szürke test Hosszú szárnyak |
Fekete test Rövid szárnyak |
|
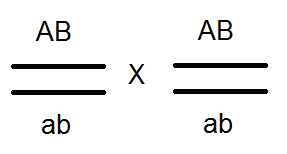
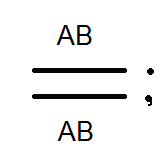
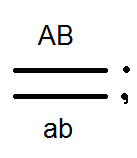
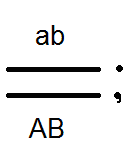
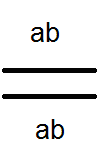
Átkelés.
A Morgan kísérleteiben az F 2-es esetek kis hányadában a legyek megjelentek új karakterkombinációkkal: a szárnyak hosszúak, a test fekete; a szárnyak rövidek és a test szürke. Ie A "feloldott" jelek. Morgan ezt elmagyarázta azzal a ténnyel, hogy a kromoszómák konjugáció során a meiosis cseréjében gének. Ennek eredményeként az egyéneket új karakterek kombinációjával kapjuk, azaz Mendel harmadik törvényének megfelelően. Morgan ezt a géncsere rekombinációt nevezte.
Később a citológusok valóban megerősítették Morgan hipotézisét, a kromoszóma-régiók cseréjét a kukoricában és a szalamanderekben. Ezt a folyamatot crossovernek hívták.
Az átkelés növeli az utódok sokféleségét a népességben.
Miért hasonlítanak a gyerekek a szüleikre? Miért gyakori bizonyos betegségek bizonyos családokban, mint például a színvakság, a polidaktikus, az ízületek hipermobilitása, a cisztás fibrózis? Miért van egy sor olyan betegség, amelyet csak a nők szenvednek, és mások csak férfiak? Ma mindannyian tudjuk, hogy az ezekre a kérdésekre adott válaszokat az öröklés, azaz a gyermek által a szülőktől kapott kromoszómákra kell keresni. És a modern tudomány ezt a tudást Thomas Hunt Morgan - amerikai genetikának köszönheti. Leírta a genetikai információ továbbításának folyamatát, és kollégáival együtt kifejlesztette az öröklési kromoszóma-elméletet (amelyet gyakran Morgan kromoszóma-elméletnek neveznek), amely a modern genetika sarokköve lett.
A felfedezés története
Helytelen lenne azt mondani, hogy Thomas Morgan volt az első, aki érdeklődött a genetikai információ átadásáért. Az első kutatók, akik megpróbálták megérteni a kromoszómák szerepét az öröklésben, tekinthetők Chistyakov, Beneden, Rabelya műveinek a XIX. Század 70-80-as éveiben.
Aztán nem voltak olyan erős mikroszkópok, hogy láthassuk a kromoszómális struktúrákat. És maga a "kromoszóma" kifejezés sem volt ott. A német tudós Heinrich Valdeyr vezette be 1888-ban.
Theodor Bowery német biológus kísérleteinek eredményeként bebizonyította, hogy a test normális fejlődéséhez normális számú kromoszómára van szüksége a fajához, és túlzott vagy hiányos súlyos fejlődési rendellenességekhez vezet. Idővel elméletét ragyogóan megerősítették. Azt mondhatjuk, hogy T. Morgan kromoszóma-elmélete pontosan a Boveri kutatásának köszönhető.
A kutatás kezdete
Az öröklési elméletre vonatkozó ismeretek általánosítása érdekében Thomas Morgan képes volt kiegészíteni és fejleszteni őket. Kísérleteinek tárgyaként a gyümölcslevelet választotta, és nem véletlenül. Ideális tárgy volt a genetikai információ átadásának tanulmányozására - csak négy kromoszómára, gyümölcsösségre, rövid élettartamra. Morgan tiszta legyek segítségével kezdte a kutatást. Hamarosan rájött, hogy a nemi sejtekben egyetlen kromoszómacsoport, azaz 2 helyett 4 volt. Morgan nevezte a női nemi kromoszómát X-nek, a férfi pedig Y-nek.
A szexuális öröklés
Morgan kromoszóma-elmélete kimutatta, hogy bizonyos jelek kapcsolódnak a szexhez. Az elülső látvány, amellyel a tudós kísérleteket végzett, általában vörös szemű színnel rendelkezik, de ennek a génnek a mutációja következtében fehér szemű egyének jelentek meg a populációban, és közöttük sokkal több férfi volt. Az X-kromoszómán lokalizálódik a legyek szemének színeért felelős gén, nem az Y-kromoszómán. Ez azt jelenti, hogy amikor egy nőt keresztezünk, egy X-kromoszómán, amelynek mutált génje van, és egy fehér szemű hím, az ilyen tulajdonságnak az utódokban való jelenlétének valószínűsége a nemhez kapcsolódik. A legegyszerűbb módja annak, hogy az ábrán látható legyen:
- P: XX "x X" Y;
- F 1: XX, XY, X "X", X "Y.
X - a női vagy férfi nemi kromoszóma fehér szeme génje nélkül; X "- kromoszóma a fehér szemek génjével.
Megfejtjük az átkelés eredményeit:
- XX "- vörös szemű nő, a korábbi szemek génjének hordozója. A második X kromoszóma jelenléte miatt ez a mutáns gén" átfedi "az egészséges és nem jelenik meg a fenotípusban.
- X "Y egy fehér szemű férfi, aki egy anyai X-kromoszómát kapott egy mutált génnel. Csak egy X-kromoszóma jelenléte miatt a mutáns tulajdonságnak nincs semmiféle blokkolása, és ez megjelenik a fenotípusban.
- X "X" - fehér szemű nő, aki a kromoszómán át örökölt egy anya és apa mutáns génjével. Egy nőben csak akkor jelenik meg a fenotípusban, ha mindkét X kromoszóma fehér szemmel rendelkezik.
Thomas Morgana elmagyarázta sok öröklés mechanizmusát genetikai betegségek. Mivel az X kromoszómán sokkal több gén van, mint az Y kromoszómán, nyilvánvaló, hogy felelős a szervezet legtöbb jellemzőjéért. Az anyából származó X kromoszómát mindkét fia és leánya átadja a test tulajdonságaiért felelős külső gének, külső jelek, betegségek. Az X-kapcsolattal együtt Y-kapcsolt örökség van. Az U-kromoszóma azonban csak férfiaknál van, így ha bármilyen mutáció bekövetkezik, csak egy férfi leszármazottja továbbíthatja azt.
Morgan öröklési kromoszóma elmélete segített megérteni a genetikai betegségek átvitelének mintáit, de a kezelésükkel kapcsolatos nehézségeket eddig nem oldották meg.
![]()
Átkelés
A kutatás folyamán Thomas Morgan Alfred Stertevant tanítványa egy átkelő jelenséget fedezett fel. Amint azt további kísérletek mutatják, az átkelésnek köszönhetően új génkombinációk jelennek meg. Ő az, aki megsérti a láncolt öröklés folyamatát.
Így T. Morgana kromoszóma-elmélete újabb fontos pozíciót kapott - közöttük átkelés történik, és gyakoriságát a gének közötti távolság határozza meg.
Főbb rendelkezések
A tudós kísérleteinek eredményeinek rendszerezéséhez bemutatjuk a Morgan kromoszóma elméletének főbb pontjait:
- A test jelei a kromoszómákban meghatározott génektől függenek.
- Az egyik kromoszóma génjeit az utódokhoz kapcsoljuk. Az ilyen kapcsolás erőssége nagyobb, annál kisebb a távolság a gének között.
- Homológ kromoszómákban megfigyelhető az átkelés jelensége.
- Ismerve egy adott kromoszóma átkelésének gyakoriságát, kiszámítható a gének közötti távolság.

A Morgan kromoszómaelmélet második pontját Morgan szabálynak is nevezik.
elismerés
A kutatás eredményeit ragyogóan érzékelték. Morgan kromoszóma-elmélete áttörés volt a huszadik század biológiájában. 1933-ban, a kromoszómák öröklődésben betöltött szerepének felfedezéséhez a tudós Nobel-díjat kapott.
Néhány évvel később Thomas Morgan megkapta a Copley érmet a genetikai kiválóságért.
Most Morgan kromoszomális öröklődési elméletét iskolákban tanulmányozzák. Sok cikket és könyvet szentelt.
Példák a nemi alapú öröklésre
Morgan kromoszóma-elmélete kimutatta, hogy a szervezet tulajdonságait az abban meghatározott gének határozzák meg. A Thomas Morgan által kapott alapvető eredmények adtak választ arra a kérdésre, hogy milyen betegségek, például hemofília, alacsony szindróma, színvakság, Brutton-betegség.
Kiderült, hogy ezeknek a betegségeknek a génjei az X kromoszómán helyezkednek el, és a nőkben ezek a betegségek sokkal ritkábban fordulnak elő, mivel az egészséges kromoszóma blokkolhatja a kromoszómát a betegséggénnel. A nők, akik ezt nem tudják, genetikai betegségek hordozói lehetnek, amelyek ezt követően gyermekekben nyilvánulnak meg.
Férfiaknál az X-kapcsolt betegségek vagy fenotípusos jelek nyilvánulnak meg, mivel nincsenek egészséges X-kromoszómák.
A genetikai betegségek családtörténetének elemzésében T. Morgan öröklési kromoszómaelméletét használják.
 A világ legszörnyűbb betegsége
A világ legszörnyűbb betegsége Teljes és hiányos dominancia
Teljes és hiányos dominancia Hogyan lehet boldogítani a barátnődet
Hogyan lehet boldogítani a barátnődet