A női nemet mindig az xx kombinációja határozza meg. Padlóbiológia
A padló genetikája
A nemeket a kromoszómákban található gének által meghatározott tulajdonságok komplexje jellemzi. Az emberi test sejtjeiben a kromoszómák párosított diploid halmazok. A kettős létszámú fajok esetében a férfiak és a nők kromoszóma-komplexe nem azonos és egy kromoszóma-pár (nemi kromoszóma) szerint különbözik. Ennek a párnak az azonos kromoszómáit X (X) -kromoszómának nevezték el, a másik nemtől - Y (play) -kromoszómától mentes - nem; a többi, amelyre nincs különbség, az autoszómák (A).
A női sejtek két azonos nemi kromoszómát tartalmaznak, amelyek XX-nek nevezhetők, férfiaknál két páratlan X és Y kromoszóma képvisel. Így egy férfi és egy nő kromoszómái csak egy kromoszómában különböznek: a nő kromoszómacsoportja 44 autoszómát + XX, férfiak - 44 autoszómát + XY.
Az emberi csírasejtek szétválasztása és érése során a haploid kromoszómákkal rendelkező ivarsejtek képződnek: a tojás általában 22 + X kromoszómát tartalmaz. Így a nőkben csak egy fajta gamete képződik (az X-kromoszómával rendelkező ivarsejtek). A férfiaknál a ivarsejtek 22 + X vagy 22 + Y kromoszómát tartalmaznak, és két fajta ivarsejt képződik (a gamete az X kromoszómával és a gamete az Y kromoszómával). Ha a megtermékenyítés során az X kromoszómájú spermiumok a tojássejtbe kerülnek, nőstény csíra képződik, és az Y kromoszómával hím.
Ezért a személy nemének meghatározása attól függ, hogy a hím csírasejtekben - a sperma, a tojás, X- vagy Y-kromoszómák megtermékenyítése - jelen van-e.
A kromoszóma-nemi meghatározás négy fő típusa van:
1. A férfi nem heterogametikus; A gaméták 50% -a X-, 50% -U - kromoszómát hordoz, például ember, emlős, dipterans, bogár, bogár (4. dia).
2. A férfi neme heterogametikus; A ivarsejtek 50% -a X-, 50% nem rendelkezik nemi kromoszómával, például szöcske, kenguru (7. dia).
3. A női nem heterogametikus; A gameták 50% -a X-, 50% gamete –Y- kromoszómát hordoz, például madarak, hüllők, farkú kétéltűek, selyemhernyó (7. dia).
4. A női nem heterogametikus; A gaméták 50% -a hordozza az X-, 50% -uk nem rendelkezik szexuális kromoszómával, például egy mol.
A nemi kromoszómákban lokalizált gének örökségét öröklésnek nevezik, a padlóhoz csatlakozik.
26. Genotípus mint integrált rendszer. A gének kölcsönhatása, a gének többszörös hatása.
Genotípus tetszik holisztikus rendszer
A gének tulajdonságai. Alapul keresek a mono- és di-hibrid kereszteződések tulajdonságainak öröklődésének példái azt a benyomást keltik, hogy a szervezet genotípusa különálló, egymástól függetlenül ható gének összegéből áll, amelyek mindegyike meghatározza saját tulajdonságának vagy tulajdonságának fejlődését. Egy ilyen tulajdonság közvetlen és egyértelmű kapcsolatának ilyen elképzelése leggyakrabban nem felel meg a valóságnak. Tény, hogy az élő szervezetek jelei és tulajdonságai hatalmas számban vannak, amelyeket két vagy több génpár határoz meg, és fordítva, az egyik gén gyakran sok jelet vezérel. Ezen túlmenően a gén hatása megváltozhat más gének és környezeti feltételek közelségével. Így az ontogenezisben nem egyéni gének működnek, hanem az egész genotípus, mint összetett rendszer, összetett kapcsolataival és összetevői közötti kölcsönhatásokkal. Ez a rendszer dinamikus: az új allélok vagy gének mutációk következtében történő megjelenése, az új kromoszómák kialakulása és még az új genomok a genotípus észrevehető változását idézik elő.
A genotípus összetételében a génaktivitás megnyilvánulásának jellege a rendszerben eltérő lehet befolyása különböző tényezők. Ez könnyen látható, ha figyelembe vesszük a gének tulajdonságait és a jelek megjelenésének jellemzőit:
A gén hatásában diszkrét, azaz más gének hatására izolálódik.
A gén specifikus a megnyilvánulásában, azaz felelős a szervezet szigorúan meghatározott jellemzőjéért vagy tulajdonságáért.
A gén fokozatosan hathat, azaz növelheti a tulajdonság megnyilvánulási fokát a domináns allélek számának növekedésével (géndózis).
Egyetlen gén befolyásolhatja a különböző tulajdonságok kialakulását - ez egy gén többszörös vagy pleiotróp hatása.
A különböző gének ugyanolyan hatást gyakorolhatnak ugyanazon tulajdonság (gyakran kvantitatív jellemzők) kialakulására - több gén vagy poligén.
A gén kölcsönhatásba léphet más génekkel, ami új jelek megjelenéséhez vezet. Az ilyen kölcsönhatás közvetett módon történik - a kontrolljaik által szintetizált reakciók termékein keresztül.
A gén hatása módosítható annak helyének megváltoztatásával a kromoszómán (pozicionáló hatás) vagy a különböző környezeti tényezők hatásával.
Az allélgének kölcsönhatása. A jelenséget, amikor több gén (allél) felelős egy tulajdonságért, nevezik gén kölcsönhatásnak. Ha ezek ugyanazon gén alléljai, akkor az ilyen kölcsönhatásokat hívják allél és különböző gének alléljainak esetében - nemallelikus.
Megkülönböztetjük az alléli kölcsönhatások főbb típusait: dominancia, hiányos dominancia, túlzott túlsúly és kodominancia.
uralom - az egyik gén két allélje közötti kölcsönhatás típusa, amikor egyikük teljesen megszünteti a másik megnyilvánulását. Ilyen jelenség a következő körülmények között lehetséges: 1) a heterozigóta állapotban lévő domináns allél olyan termékek szintézisét biztosítja, amelyek elegendőek ahhoz, hogy a domináns homozigóta állapotában ugyanolyan minőségű tulajdonságot nyújtsanak, mint a szülő formában; 2) a recesszív allél teljesen inaktív, vagy aktivitásának termékei nem lépnek kölcsönhatásba a domináns allél aktivitásának termékeivel.
Az allélgének ilyen kölcsönhatásának példái a lila dominancia szín borsóvirágok fehér, sima vetőmag alakú ráncos, sötét haj felett fény felett barna szemek kék felett embereken stb.
Hiányos dominancia vagy az öröklés közbenső jellege amikor a hibrid fenotípusa (heterozigóta) különbözik mindkét szülői homozigóta fenotípusától, azaz a tulajdonság expressziója közbenső, többé-kevésbé eltér az egyik vagy a másik szülő felé. Ennek a jelenségnek az a mechanizmusa, hogy a recesszív allél inaktív, és a domináns allél aktivitásának mértéke nem elegendő ahhoz, hogy biztosítsa a domináns jellemző megnyilvánulásának kívánt szintjét.
A hiányos dominancia példája az öröklés. szín virágok az éjszakai szépségű növényekben (3.5. ábra). Amint az a diagramból látható, a homozigóta növényeknek vörösek (AA) vagy fehér (Aa) virágok, és heterozigóta (Aa) - rózsaszín. Amikor piros virágokkal és fehér virágokkal rendelkező növényekkel haladunk át az F 1-ben, minden növénynek rózsaszín virágai vannak, azaz van az öröklés közbenső jellege. Hibridek átlépésekor a rózsaszín virágok F 2 a domináns homozigóta óta a fenotípus és a genotípus hasítása egybeesik (AA) különbözik a heterozigótáktól (Aa). Szóval, ebben a példában az éjszakai szépségű növények, amelyekre felosztották F 2 A virág színe általában a következő - 1 piros (AA): 2 rózsaszín (Aa): 1 fehér (Aa).
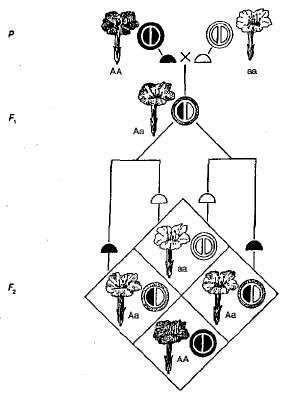
Ábra. 3.5. A virág színe öröksége éjszakai szépség hiányában.
A hiányos dominancia elterjedt. Megfigyelhető a göndör haj öröksége az emberekben, a szarvasmarha színe, a csirkék tollazatának színe, és sok más morfológiai és fiziológiai karakter a növényekben, állatokban és emberekben.
superdominance - a tulajdonság erősebb megnyilvánulása egy heterozigóta egyénben (Aa), mint bármely homozigóta (AA és aa). Feltételezzük, hogy ez a jelenség a heterózis alapját képezi (lásd 3.7. Pont).
Kodaminirvanie- mindkét allél részvétele a tulajdonság meghatározásában heterozigóta egyénben. A kodominancia szembetűnő és jól tanulmányozott példája a IV vércsoport öröklődése az emberekben (AB csoport).
Az ebben a csoportban élő emberek vörösvértestjei kétféle antigénnel rendelkeznek: antigén A (a kromoszómák egyikében található genom / \\ t az (amelyet egy másik homológ kromoszómán lokalizált genom / a határoz meg). Csak ebben az esetben mindkét allél 1 A (A a homozigóta állapot kontrollálja a II én B (a homozigóta állapotban a III. vércsoport, a B csoport). allél 1 A és én B dolgozzon egy heterozigótaban, mintha egymástól függetlenül lennének.
Öröklési példa csoportok a vér mutatja és megnyilvánul többszörös allélizmus: a gén / három különböző allél képviselhető, és vannak olyan gének, amelyek több tucat allélt tartalmaznak. Ugyanezen gén összes allélét nevezzük el több allél sorozat, ebből mindegyik diploid organizmus két allélt tartalmazhat (és csak). Ezen allélek között az allél kölcsönhatások összes felsorolt változata lehetséges.
A többszörös allélizmus jelensége a természetben gyakori. Számos több allél létezik, amelyek meghatározzák a gombákon való megtermékenyítés során a kompatibilitást, a vetőmagnövényekben történő beporzás, az állati szőr színének meghatározása stb.
A nem-gén interakciók A nem allél gén kölcsönhatásokat számos növényben és állatban írják le. Ezek a fenotípus szerint 9: 3: 4 szerinti szokatlan hasadást okozó digerozigóta utódaihoz vezetnek; 9: 6: 1; 13: 3; 12: 3: 1; 15: 1. a Mendeli általános képlet módosítása: 9: 3: 3: 1. Két, három és több nem-gén gén kölcsönhatása van. Ezek között a következő főbb típusok tartoznak: komplementaritás, episztázis és polimerek.
kiegészítő, vagy további, Ezt az interakciót nem-párhuzamos domináns géneknek nevezzük, aminek következtében mindkét szülő hiányzik egy jellemző. Például, ha két édesborsófajtát fehér virágokkal keresztezünk, az utódok lila virágokkal jelennek meg. Ha egy fajta genotípusát jelöli meg AAbb, és a másik aABB, az
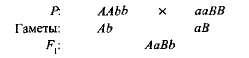
Az első generációs hibrid két domináns génnel (A és in) biokémiai alapot kapott az antocianin lila pigment előállításához, míg a gén egyenként és Egyik B gén sem szolgáltatta e pigment szintézisét. Az antocianin szintézise egy komplex lánc, amely több egymástól eltérő gén által szabályozott szekvenciális biokémiai reakciókat tartalmaz, és csak akkor, ha legalább két domináns gén van. (A-B) lila szín alakul ki. Más esetekben (aaV- és A-bb) a növény virágai fehérek (a „-” jel a genotípus formulában azt jelenti, hogy ezt a helyet mind a domináns, mind a recesszív allél foglalhatja el).
Amikor magától beporzó édes borsó növények F 1 -ban F 2 lila és fehér virágos formákba való hasítása 9: 7 arányban volt megfigyelhető. Lila virágokat találtak 9/1 6 növény, fehér - 7/16. Pennet rácsa egyértelműen mutatja a jelenség okait (3.6. Ábra).
epistasis - ez egyfajta gén kölcsönhatás, amelyben az egyik gén allélje elnyomja egy másik gén allélpárjának megnyilvánulását. gének gének megakadályozzák más gének működését episztatikus inhibitorok vagy csökkentők. Az elnyomott gént hívják hypostatic.
A fenotípus és a ciklikus osztályok számának és arányának változása szerint a Dybrid féle hasítás során F 2 vegye figyelembe az episztatikus kölcsönhatások több típusát: domináns episztázis (A\u003e B vagy B\u003e A) hasítással 12: 3: 1; recesszív episztázis (a\u003e B vagy b \u003e A), amely 9: 3: 4 stb.
polymerism nyilvánvalóvá válik abban a tényben, hogy egy jel alakult ki befolyása több gén azonos fenotípusos expresszióval. Ezeket a géneket hívják polimer. Ebben az esetben elfogadták a gének egyértelmű tulajdonságát a tulajdonság fejlődésére. Például, amikor a Pásztor zsáknövényeit háromszög alakú és ovális gyümölcsökkel (hüvelyekkel) keresztezzük az F 1-ben, háromszög alakú gyümölcsök képződnek. A saját beporzással F 2 15: 1 arányú háromszög alakú és ovális hüvelyes növényekre történő felosztást figyeltünk meg. Ez azzal magyarázható, hogy két gén egyedülállóan működik. Ezekben az esetekben azonosak. A 1 és A 2 .
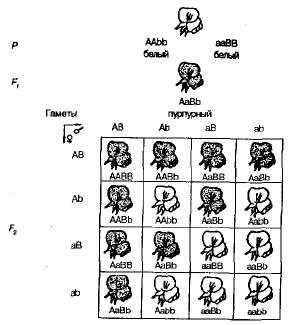
Ábra. 3.6 . Virágszín öröksége édes borsóban
Ezután minden genotípus (A 1 ,-A 2 ,-, A 1 s 2 és 2 egy 1 egy 1 A 2 -) ugyanaz a fenotípus - háromszög alakú hüvely, és csak a növények és 1 és 1 és 2 egy 2 különböző lesz - ovális hüvelyek kialakítása. Ez a helyzet nem kumulatív polimerek.
A polimer gének típus szerint működhetnek kumulatív polimerek. Minél hasonlóabb a gén a szervezet genotípusában, annál erősebb ez a tulajdonság, vagyis a gén dózisának növekedésével. (A 1 A 2 A 3 stb.) tevékenységét összefoglalják vagy összevonták. Például a búza szemek endospermájának színintenzitása arányos a különböző gének domináns alléljeivel a trihibrid kereszteződésben. A legszínezettebb szemek voltak A 1 A 1 A 2 A 2 A 3 , A 3 és gabona a 1 és 1 és 2 egy 2 és 3 és 3 nem volt pigmentje.
A halmozott polimer típusától függően számos jele öröklődik: a tejesség, a tojástermelés, a súly és a haszonállatok egyéb jelei; az ember fizikai erejének, egészségének és szellemi képességeinek számos fontos paramétere; a szemek fülének hossza; cukortartalom a cukorrépa - gyökerekben vagy a lipidekben. \\ t napraforgómag és így tovább
Így számos megfigyelés arra utal, hogy a legtöbb jel megnyilvánulása az egymással kölcsönhatásba lépő gének és környezeti feltételek hatásának az eredménye, az egyes sajátos tulajdonságok kialakulására.
Gén kölcsönhatás
A gének és a tulajdonságok közötti kapcsolat meglehetősen bonyolult. Egy szervezetben nem mindig egy gén határoz meg csak egy tulajdonságot, és fordítva, egy tulajdonságot csak egy gén határoz meg. Gyakran egy gén egyidejűleg több jel megjelentetéséhez is hozzájárulhat, és fordítva. A szervezet genotípusa nem tekinthető önálló gének egyszerű összegének, amelyek mindegyike nem érintkezik a többiekkel. A tulajdonság fenotípusos megnyilvánulása számos gén kölcsönhatásának eredménye.
Többszörös génaktivitás (pleiotropia) - egyetlen gén befolyásoló folyamatai több karakter kialakulásához.
Például az emberekben a vörös hajszínt meghatározó gén könnyebb bőrt és szeplőket okoz.
Néha a morfológiai jellemzőket meghatározó gének befolyásolják a fiziológiai funkciókat, csökkentik a vitalitást és a termékenységet, vagy végzetesek. Így a gél kék színét okozó gén csökkenti a gyümölcsösségét. A homozigóta állapotban az arakul juhokban a szürke szín domináns génje részletes, mivel az ilyen bárányok nem fejlett gyomorral rendelkeznek, és amikor a fűre váltanak, meghalnak.
Kiegészítő gén kölcsönhatás. Számos gén befolyásolhatja egyetlen tulajdonság kialakulását. A több nem-egymástól eltérő gén kölcsönhatását, ami egyetlen vonás kialakulásához vezet, komplementernek nevezzük. Például a csirkéknek négy formája van, ezek bármelyikének megnyilvánulása két pár párhuzamos gén kölcsönhatásával függ össze. A rózsás címer az egyik allél domináns génjének, a borsó alakú címernek, a másik allél domináns géneknek az eredménye. Hibridekben két domináns, nem-párhuzamos gén jelenlétében egy anya-alakú címer képződik, és minden domináns gén hiányában, azaz a génekben. recesszív homozigótákban két nem-gén gén esetében egy egyszerű fésű alakul ki.
A gének kölcsönhatásának eredménye a gyapjú színe kutyákban, egerekben, lovakban, sütőtök alakjában, édes borsóvirágok színében.
A polimer a nem-párhuzamos gének kölcsönhatása, amikor a tulajdonság fejlődésének mértéke a domináns gének teljes számától függ. Ezen elv szerint a zab és a búza szemek színezése és a bőr bőrszíne öröklődik. Például a feketékben 4 domináns gén van két pár párhuzamos génben, és egyetlen fehér bőrű emberben sem minden gén recesszív. Különböző mennyiségű domináns és recesszív gének a bőrszín különböző intenzitású mulattosainak kialakulásához vezethet: sötét és világos.
A gén kölcsönhatásnak két fő csoportja van: az allélgének kölcsönhatása és a nem allél gének közötti kölcsönhatás. Ugyanakkor meg kell érteni, hogy ez nem a gének fizikai kölcsönhatása, hanem az elsődleges és másodlagos termékek kölcsönhatása, amely egy vagy másik tulajdonságot határoz meg. A citoplazmában a fehérjék - enzimek, amelyek szintézisét gének határozzák meg, vagy az enzimek hatására képződő anyagok között fordul elő.
A következő típusú interakciók lehetségesek:
1) egy specifikus tulajdonság kialakításához két enzim kölcsönhatása szükséges, amelynek szintézisét két nem allélgén határozza meg;
2) az egyik gén részvételével szintetizált enzim teljesen elnyomja vagy inaktiválja az enzim hatását, amelyet egy másik nem-gén gén alakított ki;
3) két enzim, amelyek kialakulását két nem allélgén szabályozza, amelyek egy tulajdonságot vagy egy folyamatot befolyásolnak, hogy közös cselekvésük a tulajdonság megnyilvánulásának kialakulásához és fokozásához vezet.
Allél gén kölcsönhatás
A homológ kromoszómákban azonos (homológ) lokuszokat tartalmazó géneket allélnek nevezzük. Minden szervezetnek két allélgénje van.
Az allélgének közötti kölcsönhatások ilyen formái ismertek: teljes dominancia, hiányos dominancia, kodominancia és túladagolás.
Az interakció fő formája a teljes dominancia, amelyet először G. Mendel ír le. Lényege abban rejlik, hogy egy heterozigóta szervezetben az egyik allél megnyilvánulása dominál a másik megnyilvánulása felett. Az 1: 2: 1 genotípus szerinti felosztás teljes dominanciájával nem egyezik meg a 3: 1 fenotípusú hasítással. Az orvosi gyakorlatban a kétezer monogén örökletes betegségből majdnem félig létezik a kóros gének domináns megnyilvánulása a normál gének felett. A heterozigótákban a patológiás allél a legtöbb esetben a betegség jeleivel (domináns fenotípus) nyilvánul meg.
A hiányos dominancia az interakció egy olyan formája, amelyben a heterozigóta szervezetben (Aa) lévő domináns gén (A) nem gátolja teljesen a recesszív gént (a), aminek következtében a szülői jelek közti közbenső jelenik meg. Itt a genotípus és a fenotípus hasítása azonos és 1: 2: 1
Amikor a heterozigóta szervezetekben a kodominánsok mindegyike allélgénnel rendelkezik, a terméktől függő termék keletkezik, vagyis a két allél termékei kiderülnek. Az ilyen megnyilvánulás klasszikus példája a vércsoport rendszer, különösen az ABO rendszer, amikor az emberi eritrociták antigéneket hordoznak a felszínen, amelyeket mindkét allél szabályoz. Ezt a megnyilvánulási formát kodominancianak nevezik.
Szuperdominancia - ha a domináns gén kifejezettebb a heterozigóta állapotban, mint a homozigóta állapotban. Így Drosophilában az AA genotípusú normál élettartam; Aa - kiterjesztett trivatista élet; - halálos.
Többszörös allizmus
Minden szervezetnek csak két allélgén van. Azonban, gyakran a természetben, az allélek száma több mint két lehet, ha egyes lókuszok különböző állapotokban lehetnek. Ilyen esetekben több allél vagy több allomorfizmus mondható.
Többszörös allélt azonos betűvel jelölnek különböző indexekkel, például: A, A1, A3 ... Az allélgének ugyanazon a homológ kromoszómák területén találhatók. Mivel a kariotípusban mindig két homológ kromoszóma van, még többszörös allél esetén is, mindegyik organizmusnak ugyanakkor csak két azonos vagy különböző allélja lehet. Csak egyikük belép a reproduktív sejtbe (a homológ kromoszómák különbségével együtt). Többszörös allélok esetében az azonos tulajdonságú allélok jellemző hatása. Ezek közötti különbség csak a tulajdonság fejlődésének mértékében rejlik.
A második jellemző az, hogy a diploid szervezetek szomatikus sejtjei vagy sejtjei legfeljebb két több allélt tartalmaznak, mivel ugyanabban a kromoszómában találhatók.
Egy másik jellemzője számos allélnek tulajdonítható. A dominancia jellegéből adódóan az allélomorf karakterek egymás után kerülnek sorba: gyakrabban a normális, változatlan tulajdonság dominál a többiekben, a sorozat második génje recesszív az elsőhez képest, de uralja a következőt, stb. A többszörös allélok egy személyben való megnyilvánulásának egyik példája az ABO rendszer vércsoportjai.
A többszörös allizmus fontos biológiai és gyakorlati jelentőséggel bír, mivel növeli a kombinációs variabilitást, különösen a genotípusos változást.
A nem allél gének kölcsönhatása
Számos olyan eset van, amikor egy tulajdonságot vagy tulajdonságokat két vagy több egymástól elválaszthatatlan gén határoz meg. Bár itt az interakció feltételes, mert nem a gének kölcsönhatásba lépnek, hanem az általuk irányított termékeket. Ebben az esetben eltérés van a mendeli hasítási mintáktól.
A gén kölcsönhatás négy fő típusa van: komplementaritás, episztázis, polimerek és módosító hatás (pleiotropia).
A komplementaritás olyan fajta kölcsönhatás a nem-gén gének között, amikor az egyik domináns gén egy másik nem-domináns domináns gén hatását kiegészíti, és együtt határozzák meg a szülőkben hiányzó új tulajdonságot. Ezenkívül a megfelelő tulajdonság csak a két nem allél gén jelenlétében alakul ki. Például a kén a gyapjú színe két gén (A és B) által szabályozott egerekben. Az A gén meghatározza a pigment szintézist, azonban mind a homozigóták (AA), mind a heterozigóták (Aa) albínó. Egy másik B-gén pigment-klasztereket biztosít a haj alapjain és csúcsán. A diheterozigóta keresztezése (AaBb x AaBb) a hibridek szétválasztásához vezet 9: 3: 4 arányban. A kiegészítő kölcsönhatások numerikus arányai 9: 7 lehetnek; 9: 6: 1 (Mendeleev-hasítás módosítása).
Az emberben a gének komplementer kölcsönhatása lehet egy védőfehérje, interferon szintézise. A szervezetben való kialakulása a különböző kromoszómákban található két nem allél gén komplementer kölcsönhatásával függ össze.
Az episztázis egy olyan nem-gén gén kölcsönhatása, amelyben az egyik gén egy másik nem-gén gén hatását elnyomja. Mind a domináns, mind a recesszív gének depressziót okozhatnak (A\u003e B, A\u003e B, B\u003e A, B\u003e A), és ennek függvényében az episztázis dominál és recesszív. A túlnyomó gént gátlónak vagy szupresszornak nevezik. Az inhibitor gének általában nem határozzák meg egy adott tulajdonság fejlődését, hanem csak egy másik gén hatását gátolják.
A gén, amelynek hatását elnyomják, hypostatikusnak hívják. Episztatikus gén kölcsönhatás esetén az F2 fenotípus-hasítása 13: 3; 12: 3: 1 vagy 9: 3: 4, stb. A sütőtökfajták színe, a lovak színe az ilyen típusú interakció alapján történik.
Paul - ez a test morfológiai, fiziológiai, biokémiai, viselkedési és egyéb jeleinek halmaza, ami reprodukciót okoz.
Azok a jelek, amelyeken a különböző nemek egyének különböznek, az elsődleges és a másodlagos. Az elsődleges szervek olyan szervek, amelyek biztosítják a gameták és a termékenyítés (gonádok, nemi szervek, szervek) kialakulását. Ezek olyan külső és belső nemi szervek, amelyek embriogenezisbe kerülnek. Másodlagos - nem vesz részt szexuális reprodukcióban. A nemi hormonok hatására alakul ki és pubertáskor jelentkezik (emberben 12-15 év). Ezek az izom-csontrendszer, a szubkután zsírszövet, a haj, a hangszóró, a viselkedési jellemzők fejlődésének jellemzői; madarak, ének, tollazat stb.
A nemhez kapcsolódó egyének jelei három kategóriába sorolhatók:
1) a padlóra korlátozódik
2) a padló vezérlése
3) a padlóhoz csatlakoztatva.
Az előbbiek kialakulását a két nem autoszómájában található gének okozzák, de csak egy nemben jelentkeznek. Így a tojásrakó gének jelen vannak csirkékben és kakasokban, de csak csirkékben jelennek meg. Hasonlóképpen a szarvasmarhák zsír- és tejtermelésének génjei is viselkednek. Ezt a jelenséget a megfelelő nemi hormonok expozíciójával összefüggésben figyelték meg.
A második példa a hornedness megnyilvánulása a hím tehenekben, a komolost-ban. Emberekben: kopaszság férfiaknál, köszvény - 80% férfiaknál és 12% nőknél.
A nemi kromoszómák génjei által vezérelt jeleket nemi összefüggésnek nevezik. Körülbelül 200 ilyen jel van, az X kromoszómával összefüggő színvakság, hemofília; Y-kromoszóma - hypertrichosis, ichtyosis.
A VÍZI MEGHATÁROZÁS TÍPUSA.
1. Progamous - megtermékenyítés előtt. A nemi kromoszómák aránya ebben az esetben nem játszik szerepet diploid oociták. (Egyes férgek, rotiferek - nőstények nagy oocitákból, kis oocitákból származó férfiakból származnak).
2. Egyedülálló - genetikai meghatározás a szex során a megtermékenyítés során, amely a nemi kromoszómák kombinációjának természetétől vagy a nemi kromoszómák és az autoszomák arányától függ.
3. Epigámiás - a külső környezet hatása alatt (bonelia féreg).
A VILÁG MÓDOSÍTÁSÁNAK MÓDOSÍTÁSA
┌─────────────────────────┬───────────────────┬──────────────────┐
│ Kromoszóma típusok │ genotípusok ete gamete típusok │
│ nemi meghatározás ├─────────┬─────────┼────────┬─────────┤
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
│ hím heterogame │ │
Ork Gólya rovarok │ │ │ │ │
│ (hibákat Protenor, bogarak, │ │ │ │ ugs
│pauki, szöcske) │ ХО │ ХХ │ Х, О │ Х │
Ros Drosophilus │ XY │ XX │ X, Y X │
│ gerincesek │ │ │ │ │
│ (emlősök, ember) │ XY │ XX │ X, Y │ X │
│ │ │ │ │ │
│ női heterogamma │ │
Irds Madarak. hal, pillangók, │ │ │ │ │
│ коп │ │ │ │
│ vízi. │ XX │ XY │ X │ X, Y │
IceÁr és egyéb erőteljes │ │ │ │ │
Éjszaka │ XX │ XO │ X │ X, O
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
A syngamic definíció típusa a kromoszóma meghatározása genetikai kontrollokkal rendelkező gének. Felelős a nemi kromoszóma nemében. Egy normál férfi gamete hordozza az X vagy az Y kromoszómát és az összes tojást - az X kromoszómát. Normál kromoszóma-eltérés esetén a meiosis során normális tojásokat és spermát képeznek az X és Y kromoszómák szokásos halmazával, a zigóta nemét a XX. És XY-ivarok aránya határozza meg (homogetikus és heterogametikus).
Correns nemi kromoszóma elmélete (1907) az, hogy a szexet a nemi kromoszómák kombinációja határozza meg a megtermékenyítés során. A következő típusú kromoszóma-nemi meghatározást különböztetjük meg: XY, XO, ZW, ZO.
A mitózis vagy meiózis lefolyásának megsértése esetén ginandomorfok alakulhatnak ki. Az ilyen személyek különböző sejtjeiben lévő nemi kromoszómák tartalma eltérő lehet (mozaik). Drosophila repülés: XX és XO, a XX és XY emberben, amelyekkel kapcsolatban a különböző testfájdalmak megfelelő szexuális jelei lehetnek. Lehet, hogy más mozaikszerű esetek is vannak: XX / XXX, XY / XXX; XO / XXY és mások
Ha a nemi kromoszómák nem szakadnak meg, a humán zigótákban 12 szexuális kromoszóma lehetséges kombinációja lehet, ami az emberi kromoszóma-aberrációk oka.
│ X │ XX │ O
─────┼────────┼─────────┼────────
X │ XX │ XXX │ XO
Y │ XY │ XXY │ YO
XY Y XXY │ XXXY │ XYO
O │ XO │ │
A nemi kromoszómák mioziszisztizálása nélkül a XX. És az O-gamétákat nőstényekben alakítják ki. és XY és O férfiaknál. A termékenyítéssel való részvételük során a zigótákat a nemi kromoszómák szokatlan kombinációjával képezik. Emberekben az ilyen rendellenességek 600-700 újszülöttnél előfordulnak. A Zygote Yo korai szakaszban hal meg; XXX, XXY, XO egyének életképesek, és nemük függ az „Y” kromoszóma jelenlététől vagy hiányától, amely bármilyen számú X kromoszómával szabályozza a férfi nemi jelek kialakulását, a fejlődést és serkenti a herék kialakulását. Az X kromoszómák feleslege alkotmányos anomáliákat és intelligenciahibákat okoz. A természetben azonban vannak olyan egyének, amelyekben az „Y” kromoszóma genetikailag inert, és nincs különösebb befolyása a szex meghatározására.
A CW típusú Drosophila példányok férfiak, de meddőek (1916, C. Bridges) voltak, és a XXY egyének normális termékeny nőstények.
A nemek egyensúlyi elmélete (Bridges, 1922). Megvizsgálta a nemi szervek és az autoszomák arányát
A 2n kromoszómákat tartalmazó normál nőknél az autoszomák és az X kromoszómák aránya 1: 2n = 2A + 2X (2X: 2A = 1 - normális nő), 1, 5 - a nőstény fölött: 2A + 3X (3X: 2A = 1, 5 - eredménytelen). Férfiaknál az arány 0, 5 2n = 2A + XY (X: 2A = 0, 5). Az egyének csökkenésével a férfi 3A + XY (X: 3A = 0, 33 - kopár) - szupermen marad. Az 1 és 0, 5 közötti együttható értéke megfelel a nemek közötti közbenső fenotípusnak: 3A + 2X (2X: 3A = 0, 66 - mindkét nem jelei meddőek).
Így az egyensúlyelmélet lényege, hogy nem csak a nemi kromoszómák, hanem az autoszómák is részt vesznek a nemek meghatározásában. Az autoszómák egy haploid halmaza megmondja az egyes hím tulajdonságait. Ebben az esetben a padlót az autoszómák és a nemi kromoszómák száma (egyensúly) aránya határozza meg.
A méhekben szintén megtalálható a ploidiás nemi meghatározás. A nőstények diploidok, a férfiak haploidok, mivel a nem szaporodott tojásokból származó partenogenetikai fejlődést.
A FLOOR DONFERENCIÁJA ONTOGENESIS-en
A korai embriókban a gonádok kezdetei (az 5. vagy a 6. hétig) nem különböznek egymástól a nemek között, és a külső rétegből állnak - a kéreg kéregéből és a belső rétegből - nem tartalmaz csíravonalakat. Az elsődleges csíravonal-sejteket az embriófejlődés harmadik hetében találták meg a sárgabarack ectodermében. Ezután a kemotaktikus jelek hatására migrálnak a gonádokra. Ez a migráció független a nemtől. A gonadok rudimentumai fejlődhetnek a petefészkekben vagy a herékben. A 8. héten megfigyelhető a gonád differenciálódás: a 36. napon a herék androgéneket (tesztoszteron) szekretálnak, ami meghatározza a férfi nemi fejlődését.
A nemi mirigyeket tartalmazó szervezetekben a szexuális jellemzők kialakulásának genetikai ellenőrzése történik.
A nemi megkülönböztetést a nemi kromoszómák genetikai halmaza programozza, amely a női és a férfi ivarsejtek összefolyásakor keletkezik. Az embrió genetikai nemét a XX vagy XY nemi kromoszómák határozzák meg.
A nemi fejlődés irányát az Y kromoszóma jelenléte határozza meg. Általában az X-kromoszómák represszor gént (Tfm, a herék feminizációs gént) tartalmaznak, amely megakadályozza a férfi típus kialakulását. A gén normál allélja meghatározza az androgének fehérje receptorának szintézisét, amelyet mindkét nemben szintetizálnak. A hím fenotípus fejlődése az Y-kromoszóma géntől, a HY-antigéntől (az egérben 1955-ben leírva, a HA géntől) függ. A primer férfi csíravonal sejtjeit választja ki. Az Y 5NA 0 felelős a tesztoszteron (férfi nemi hormon) előállításáért. Amint ezek a sejtek bejutnak a gonádok kezdetébe, a herék differenciálódása megkezdődik. A HY receptorok mindkét típusú gonad sejtek felületén vannak jelen (a teheneknél a nemi fejlődéstől való eltérés). Úgy vélték, hogy a hím fenotípust a teljes férfi kromoszóma határozza meg. De 1990-ben a gént felfedezték (Y szexuális régió), amely az Y kromoszóma kariotípusában lokalizálódott. Ennek hiányában az XY genotípus női fenotípust ad.
A GONADOK FEJLESZTÉSE
│ ┌┬┬┬┐ │ Gonad csíra biszexuális
│ ├┼┼┼┤ │ talnaya
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
ha a genotípus X │ │, ha a genotípus XY
(7-8 hét) ┴──┴──┐ ┌───┴───┐ (6. hét)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
kortikális │ └┴┘ │ │└┴┴┴┴┴┘│ medulláris réteg
└─────┘ └───────┘ réteg
(a gén a női nemet határozza meg) (a gén a férfi nemét határozza meg)
Az emberekben a specifikus női hormonok szabályozzák a
ezek a jelek észrevehetővé válnak, ezek intersticiális termékek
7-8 hét végén. A petefészek vagy a magsejtek 20. szövetén
a sejt primer csírasejtjei (Leydig sejtek). Ők alkotnak
hormonális szex (tesztoszteron,
és az ösztradiolra differenciált). 10-12 hét
az oogonyev második hónapjának végéig. belső szexuális szervezetek
A harmadik hónap végéig, a 12. héten, bizonyos fokig
az embrióban az androgének szintjén detektálnak gonad embriókat
oociták (MI profil). Elkezdődik a differenciál masculinizáció (
petefészek-ciszták 7 hónapos korukig (középen) és befejeződnek
a hetedik hétig a petefészekben a 9. hónapig is megolvad.
200-400 ezer petesejt II. A pubertás, az est szintje
a rogen növekszik és befolyásolja
csontvázszerkezet, valamint
androgének (a. \\ t
női és férfi típusú).
A gonádok meghatározzák az elsődleges szexuális jellemzők fejlődését és a másodlagos jellemzőket. A nemi mirigyek hormonokat (ösztradiolokat, androgéneket) szekretálnak, amelyek az endokrin mirigyek hormonjaival együtt szabályozzák a nemi differenciálódás útját. A hormonok szintjét viszont gének szabályozzák.
Így a szexuális megkülönböztetés folyamata:
1) genetikai ellenőrzés;
2) a hormonok szabályozási funkciói.
Van egy elmélet a hormonok hatásáról a gének szabályozói tényezőként. Csak specifikus célsejtekre hatnak. Egy speciális fehérje keletkezik a sejtben - egy receptor, amely a hormonhoz kötődik, és ugyanakkor változik, majd megszerzi a kromoszómákban lévő egy vagy több gén munkáját kiváltó tulajdonságokat. Amikor a glükokortikoidok a méhsejtekre hatnak, az RNS és a fehérjék szintézise megváltozik (lásd az ábrát).
A fehérje receptorok és hormonok képződését gének szabályozzák. Ellenőrzés megsértése esetén anomáliák lehetségesek, amelyek példája a Morris-szindróma. Méhsejt-feminizáció (Morris-szindróma) (Morris, 1953): a tesztoszteron receptorok nincsenek jelen ebben a betegségben szenvedő egyénekben. Az androgéneket normál mennyiségben választják ki. Maurice-szindrómában az embriogenezis alapja annak, hogy a herék hímhormonokat termeljenek. Az ilyen embriók azonban nem képeznek receptorfehérjét (recesszív génmutáció), amely biztosítja a fejlődő szervek sejtjeinek érzékenységét a férfi nemi hormonhoz. Ennek következtében a férfi típusú fejlődés megszűnik, és megjelenik a női fenotípus. Kivételes esetekben az ilyen hibákat a megfelelő hormonok bevezetésével lehet korrigálni.
Így az eredeti genetikai biszexualitás alapja a nemek újradefiniálásának. A férfi magzat megszerzi a női nemi jellemzőket. Férfi kariotípus, férfi gonád, női fenotípus. A test aránya nő, emlőmirigyek, rövidített hüvely, herék - a labia majorában, a nyaki csatornában, a hasüregben.
A TÉR ELLENŐRZÉSE
A hormonok expozíciója vagy a célsejt receptorainak patológiája következtében a nemi újradefiniálás történhet (Maurice-szindróma, a nemi mirigyek eltávolítása kakasokban).
A természetben számos tényező gyengíti a nemek fejlődését szabályozó gének hatását. Például az emberben az egyén gonádjaiban a herék és a petefészkek (a herék és a petefészek kifejlődnek) egyformán fejtik ki a hermafroditizmust - az interszexualitás jelenségét.
A klinikai adatok alapján 3 intersex-típus létezik:
1) valódi hermafroditizmus: a két nemű csírasejtek jelenléte;
2) férfi pszeudohermaphroditizmus: csak herék (herék, herék), női fenotípus;
3) női pszeudohermaphroditizmus: csak petefészek, a férfi fenotípus.
Ez a besorolás nem egyezik meg a citogenetikai bázisokkal, hiszen az emberben 46 XY van lehetőség.
Szex arány
Az elsődleges nemek aránya (a megtermékenyítés idején közel 1: 1 legyen, mivel a nemi kromoszómák találkozója egyaránt valószínű). Emberi vizsgálat során azt találtuk, hogy 100-160 hím alakul ki 100 női zygótára. Az Y kromoszómát tartalmazó spermiumok könnyebbek, mozgékonyabbak és nagy negatív töltéssel rendelkeznek (a tojás pozitív töltésű). Ezért az Y-tartalmú spermium gyakrabban termékenyíti a tojást.
Másodlagos - 100-105 fiú 100 született. Nőstények, férfi embriók idegen fehérjék életképessége. 20 éves korig 100 lány 100 gyermeke van.
Tercier - 50 éves korig 85 nő 100 nő esetében, és 85 évesen 50 nő 100 nőnként. A nőstény szervezet jobban alkalmazkodik, ami más okokkal magyarázható a női szervezet mozaikságával a nemi kromoszómákon.
M. Loyon hipotézise a nemi kromoszómákról szóló női mozaikizmusról.
1949-ben M. Borr és C. Bertrand megállapították, hogy a nők idegsejtjeinek magjaiban intenzíven színes kromatin található. A sejtek sejtjeiben nem észlelik azt. Ezt a csomót szexkromatinnak nevezik (Borra teste), és egy inaktivált X kromoszómát képvisel.
A női embrió fejlődésének kezdetén mindkét X-kromoszóma funkcionál, azaz kétszer annyi, mint a férfiak. Ez magyarázhatja a női zygóták nagyobb életképességét.
1962-ben Lyon hipotézist tett egy X kromoszóma inaktiválására a női emlős testben. A női embrióban mindkét kromoszóma az embrió fejlődésének 16. napjáig működik. A 16. napon egy kromoszóma inaktiválódik a nemi kromatin képződésével. Ez az eljárás véletlenszerű, így az aktív sejtek kb. 1/2 fenntartja az anyai X kromoszómát X 5M 0, és az apa inaktiválódik. Másokban az apa aktív (X 5O 0), és az anya inaktiválódik. Az újraaktiválás nem következik be. Az anyai és apai X-kromoszómák allél, de nem teljesen azonos géneket tartalmaznak, azaz a domináns allél egy kromoszómában található, a másik recesszív. További gének birtoklása kiterjeszti a test adaptív képességeit.
A SEXUÁLIS DIFFERENCIÁCIÓ szexuális DIAGRAM szintjei
különbségtétel
┌─┐ ┌┴┐
gamed X └┬┘ └─┘ Y
petesejt spermium
┌───────────────────┐
genetikai │hromosomy XX és XY │
└───────────────────┘
┌─────────┴─────────┐
gonád │ undefined differenciált │
│── Az embrió gonádja
│ └───────────────────┘
Y │ H Y antigén
Az ┌───────┴───────┐ ┐─────────────┐ időszakban
genitális │ │ │ │1 genitális │
érés │ │ │ ┌─│ jelek
│ │ │ │ └────────────┘
Ero ┌─────┴─────┐┐ ут in utero
│ │gondad │ │gunad ├────┤ 7-32 hét
Aries │ petefészek es herék hormonok andro
│ └───────────┘ └───────────┘┘ │ │ sejt gének
Yd │ │ Leydinga
Erty │ pubertás │ │ ┌────────────┐
└───────────────────────────────┘ │ differenciálás
hormonális αип hipofízis hormonok, férfi és női │ ideg ideg │
│ hormonok: androgének (tesztoszteron), estro-путей─│nyh módok │
└─ gének (progisteron) │ nemek │
┌──────────────┴───────────────┐ └────────────┘
fenotípusos │ Másodlagos szexuális tulajdonságok признаки
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
pszichológiai │ szexuális és viselkedési kapcsolat │
└───────────────────────────────────────────────────┘
A női test jobban ellenáll a hidegnek, az ionizáló sugárzásnak, az érzelmi túlterhelésnek (a nők gyakrabban sírnak, az aktív aminok könnyekkel kiemelkednek, ezért csökken a vérnyomás).
Ha a hipotézis korlátozás nélkül működött, nem lenne fenotípusos különbség az egészséges nők és a két X-kromoszóma és az X 4 0-os vagy XY / XXYY-es betegek között. Nyilvánvaló, hogy az X-kromoszóma nincs teljesen inaktiválva.
A nemek arányának szabályozása.
Ha a spermát állandó elektromos mezőbe helyezi, az X - és Y - kromoszómák elkülönülnek. Állatokban használják. Lehetőség van a kívánt nemi leszármazottak 80% -ának megszerzésére.
A férfiakhoz vagy nőkhöz való tartozás tudatossága a nemi mentális észlelés szerves része, a mentális paraméterek. Transzszexualizmus - pszichológiai hermafroditizmus., Az egyén szexuális identitásának megsértésének jelensége. Az ellenkező nemhez tartozó emberek rögeszméi tudatossága az ókorban ismert. Tehát, Herodotus, a "szkíta betegség" rejtélyében egy olyan férfit ír le, aki nemcsak női ruhát viselt, hanem elfogadta az ellenkező nemű karakterjellemzőit. Néhány császár hajlamos volt álcázni: például Caligula. J. D ”Ark.
Az anatómiai és mentális paraméterek közötti eltérés a belső környezet és a társadalmi konfliktus. A férfi testben él a női lélek, és fordítva. Készítsen sebészeti beavatkozást a padlóra. Ahhoz, hogy egy embert nővé alakítson, 1 műveletre van szükség: a hüvely a pénisz és a scrotum bőréből képződik. Egy nőtől, hogy nehezebbé váljon az ember: 3-4 művelet (az emlőmirigy eltávolítása, egy tag kialakítása). Ezek az emberek örök gyötrelemre ítélnek: hormonokat isznak, nincsenek gyerekeik. Az emberi reprodukciós és családtervezési intézet Moszkvában.
A szexuális megkülönböztetés magában foglalja az elsődleges (gonad) és a másodlagos szexuális jellemzők kialakulását.
A legtöbb állatban a nemek között a műtrágyázás idején genetikailag határozzák meg.
Az X-kromoszóma hordozza a normális feminizációs (Tfm) gént
amelynek allélja meghatározza az androgén fehérje receptor szintézisét, \\ t
amelyek a nők és a férfiak egyaránt szintetizálódnak.
──────┐ - minden kromoszóma nem funkcionális Tfm - jelentése
│ A Keat csalás fejfájást fejt ki.
┐────┤ └┴┴┘ ├──────┐┐ XY - tartalmazza az I 5HA-gén 0 gént, amely felelős
Antig └──────┘ │ az antigén szintézise, amely meghatározza
│ M │ férfi = felelős a gyártásért
│ │ tesztoszteron.
┌──┴──┐ ────┴───┐ A gonádok differenciálódását 7-10
Life │ │ élethét. A 10. héten a szex lehet op
│ │ │ │ meghatározni a kromoszómák halmazát.
└─────┘ └───────┘
A fejlődés irányát az Y kromoszóma jelenléte határozza meg. Általában a XX kromoszómák represszor gént tartalmaznak, amely megakadályozza a férfi típus kialakulását. Ez a fejlődés a HY (ON) antigéntől függ, az I 5HA gén felelős a tesztoszteron előállításáért. Számos fajban megtalálható (az egérben leírták 1955-ben, Eychwald, Silmser). A primer férfi csíravonal sejtjeit választja ki.
Általában a nemi kromoszómák kombinációja esetén a női gének dominálnak a női nemet meghatározó génekben, a férfiaknál pedig a férfi neme dominál.
A nemi szerveket a Müller és a Wolf csövek alkotják, amelyek az elsődleges veséből származnak. A nőknél a Müller csatornái petesejtekké és méhekké fejlődnek, és a wolfa csövek atrófia. A férfiaknál a farkascsatornák a szemes csatornákra és a magfészekké fejlődnek. A szteroid hormonok tesztoszteron és dihidrotesztoszteron szintetizálódnak az embrionális herékben az anya hormonjának (chorion gonadotropin) hatása alatt. Ezek a hormonok a biopotens rügyekre hatnak.
külső és belső nemi szervek: farkascsatornák, Mullerovy-csatornák és urogenitális sinus. Egy normális férfi szervezet fejlődik, ha ezek az elemek működnek. Ennek hiányában női nemi jellemzők alakulnak ki. A hím fenotípus férfival való hiányos fejlődése
genotípus (férfi pszeudohermaphroditizmus).
Modern reprodukciós stratégia:
- mesterséges megtermékenyítés;
- in vitro trágyázás;
- az embrió mesterséges növekedése és átadása a méhbe;
- helyettesítő anya.
A legtöbb állat kétfajta szervezet. A nemek a jellemzők és struktúrák együttesének tekinthetők, amelyek utat biztosítanak az utódok reprodukálására és örökletes információk továbbítására. A nemet a leggyakrabban a megtermékenyítés idején határozzák meg, azaz a cigóta kariotípusa a fő meghatározásában játszik szerepet a nemek meghatározásában. Az egyes organizmusok kariotípusa olyan kromoszómákat tartalmaz, amelyek mindkét nemben azonosak - autoszómák és kromoszómák, amelyekben a női és férfi nemek különböznek egymástól - nemi kromoszómák. Emberben a "női" nemi kromoszómák két X kromoszóma. A gameták kialakulása során minden tojás kap egy X-kromoszómát. Az X-kromoszómát hordozó azonos típusú nemzetségek nemét nevezik homogametikusnak. Emberben a női nem homogametikus. A "férfi" nemi kromoszómák az X kromoszóma és az Y kromoszóma. A gameták kialakulása során a spermiumok fele megkapja az X kromoszómát, a másik felét az Y kromoszóma. A különböző típusú ivarsejtű padlót heterogetikusnak nevezik. Emberben a férfi nem heterogametikus. Ha két X-kromoszómát hordozó zigót képződik, akkor egy női test alakul ki belőle, ha az X-kromoszóma és az Y-kromoszóma hím.
Állatokban az alábbiak különböztethetők meg négyféle kromoszóma-nemi meghatározás.
1. A női nem homogametikus (XX), a férfi nem heterogametikus (XY) (emlősök, különösen az ember, gyümölcsfenyő).
A kromoszómális nemi meghatározás genetikai rendszere az emberekben:
A kromoszómális nemi meghatározás genetikai rendszere Drosophilában:
2. A női nem homogametikus (XX), a férfi nem heterogametikus (X0) (orthoptera).
A sivatagban a kromoszóma-nemi meghatározás genetikai rendszere:
3. A női nem heterogametikus (XY), a férfi nem homogametikus (XX) (madarak, hüllők).
A kromoszóma nemi meghatározásának genetikai rendszere egy galambban:
Yandex.DirectAll hirdetések
4. A női nem heterogametikus (X0), a férfi nem homogametikus (XX) (bizonyos rovarok).
Szex meghatározás emberben az XY mechanizmussal történik. Ugyanakkor a heterogetikus szex hím, homogametikus - nő. A nemek közötti egyenlőség meghatározása három szakaszra oszlik: kromoszóma, gonád és fenotípus.
Két alapszabály az emlősök nemének meghatározására
A klasszikus embriogenetikai vizsgálatok két szabályt állapítottak meg az emlősök nemének meghatározására. Ezek közül az elsőt az 1960-as években Alfred Jost fogalmazta meg olyan kísérletek alapján, amelyek a korai nyúl embriókban a jövőbeli gonádok csíráinak (gonadális henger) eltávolítására irányultak: a görgők eltávolítása a gonád képződése előtt az összes embrió nőstény fejlődéséhez vezetett. Javasolták, hogy a magzatok masculinizációjáért felelős hím effektorhormon tesztoszteron gonádjai szekretálódjanak, és egy anti-Muller hormon (MIS) második effektorjának jelenlétét, amely közvetlenül ellenőrzi az ilyen anatómiai transzformációkat. A megfigyelések eredményeit szabályként fogalmazták meg: a herékben vagy petefészekben kialakuló gonádok fejlődése meghatározza az embrió későbbi szexuális differenciálódását.
1959-ig az X kromoszómák számát feltételezték az emlősökben a szexet leginkább befolyásoló tényezőnek. Azonban egyetlen X-kromoszómával rendelkező, nőstényekként fejlődő szervezetek, valamint egy Y-kromoszómával és többszörös X-kromoszómával rendelkező férfiaként kifejlődött személyek kénytelenek voltak elhagyni az ilyen fogalmakat. Az emlősök nemének meghatározására vonatkozó második szabályt megfogalmazták: Az Y-kromoszóma a férfiak nemének meghatározásához szükséges genetikai információt hordozza.
A fenti két szabály kombinációját néha növekedési elvnek nevezik: Az Y-kromoszóma jelenlétével vagy hiányával kapcsolatos kromoszómális szex meghatározza az embrió gonád differenciálódását, amely viszont a szervezet fenotípusos nemét szabályozza. A szex meghatározására szolgáló ilyen mechanizmust genetikának nevezzük. GSD) és ellenezzék a környezeti tényezők (Eng. ESD) vagy a nemi kromoszómák és az autoszomák aránya (eng. CSD).
Hormonális nemi meghatározás
A nemi definíciót batonként lehet ábrázolni, amelyet a kromoszóma-mechanizmus megkülönböztetetlen gonádoknak továbbít, amelyek férfi vagy női nemi szervekké fejlődnek. A nemi kromoszómák szerepének tanulmányozása során a gonadok fejlődésében kimutatták, hogy az Y kromoszóma jelenléte vagy hiánya meghatározó az emberekben. Az Y-kromoszóma hiányában a petefészek gonád-differenciálódása következik be, és egy nő alakul ki. Az Y kromoszóma jelenlétében a férfi rendszer fejlődik. Nyilvánvaló, hogy az Y kromoszóma olyan anyagot termel, amely stimulálja a herék differenciálódását. "Úgy tűnik, hogy a természet alapvető terve az volt, hogy egy nőt hozzon létre, és hogy az Y-kromoszóma hozzáadása variáció-embert eredményez." A relé következő lépése folytatja azokat a hormonokat, amelyek meghatározzák a magzat szexuális differenciálódásának folyamatát és anatómiai fejlődését. Születéskor a program első része véget ér. A születés után a baton olyan környezeti tényezőkhöz megy át, amelyek a nemek kialakulását fejezik be - általában, de nem mindig, a genetikai nemek szerint. A nemek közötti egyenlőség meghatározása egy összetett, többlépéses folyamat, amely az emberben a biológiai mellett a pszichoszociális tényezőktől is függ. Ez transzszexuális, heteroszexuális, biszexuális vagy homoszexuális viselkedéshez és életmódhoz vezethet.
A gonadák nemi meghatározásának fiziológiai alapja
A nemi meghatározás mechanizmusának fiziológiai alapja a magzati emlősgonádok biszexualitása. Ilyen progonádokon egyidejűleg jelen vannak a Mullerov-csatorna és a Wolf-csatorna - a nők és a férfiak genitális traktusainak baktériumai. A szex elsődleges meghatározása a Sertoli sejtben lévő speciális sejtvonalak megjelenésével kezdődik. Az utóbbiban szintetizálható a Zhost által előre jelzett Müller-ellenes hormon, amely közvetlenül vagy közvetve gátolja a Müller-csatorna fejlődését - a jövőbeni petefészek és a méh csírája.
Genetikai mechanizmus a nemek meghatározására
Emberi Y kromoszóma, jelezve a SRY gén helyét
1987-ben David Page és munkatársai egy XX-es embert vizsgáltak, aki az Y-kromoszóma 280 ezer nukleotidjának egy meghatározott részét örökölte, és egy XY nőt, aki törli ezt a régiót a kromoszómák közötti szakaszok cseréje eredményeként. Ez a fragmens az összes valódi Eutheria állat Y kromoszómájában van jelen, és 100 ezer bázispár távolságra helyezkedik el a ZFY gén pszeudoautoszómális régiójának határainál, 140 ezer bázispár hosszúságú.
A ZFY homológ - a ZFX gén megtalálható az X kromoszómán, és a ZFX nincs inaktiválva. Mind a ZFX, mind a ZFY faktorok cink-ujj motívumokat tartalmazó transzkripciós faktorokat kódolnak DNS-kötő aktivitással. A szexuális inverzióval rendelkező Y-kromoszómák specifikus szekvenciáinak további részletes elemzése a keresést 35 kbp méretű területre korlátozta, és egy olyan gén felfedezéséhez vezetett, amely a klasszikus angol igazi egyenértékének tekinthető. Testis meghatározó tényező. Ezt a gént SRY-nek hívják (eng. Szexmeghatározás Y régió gén).
SRY a nemi meghatározás területén található, és tartalmaz egy konzervatív domént (HMG-box), amely egy 80 aminosavmaradékot tartalmazó fehérjét kódol. A SRY gén aktivitását a herék differenciálódási periódusának kezdete előtt - az egér embrionális fejlődésének 10-12. Napja előtt - észleljük, és legalábbis ebben a szakaszban nem függ a csírasejtek jelenlététől. Az adott gén HMG-dobozában XY-nőkben a specifikus pontmutációk vagy deléciók a szexuális inverzióhoz vezetnek. Az ezt a gént tartalmazó 14 kbp-os DNS-fragmens áthelyezése a szomszédos régiókkal egy mikrohullámú homogametikus egyéni termékenyített tojásba, amelynek eredményeként a XX.
SRY gén funkciói
A SRY-gén HMG-boxja által kódolt domén specifikusan kötődik a DNS-hez, és kanyarodik. A SRY fehérje vagy a HMG domént tartalmazó homológok által okozott DNS-hajlítás jelentős távolságra mechanikusan továbbítható, és fontos szerepet játszik a transzkripció, a replikáció és a rekombináció szabályozásában. A DNS azon régiója, ahol a SRY lokalizálódik, két olyan gént kódol, amelyek a férfi típusú elsődleges gonád differenciálódásban részt vevő kulcs enzimeket kódolnak: a P450 aromatáz gén, amely szabályozza a tesztoszteron ösztradiolra és a Miller csatorna kialakulásának gátlójává történő átalakulását, ami ellentétes fejlődést okoz és elősegíti a herék differenciálódását .
A SRY géntermék szintén részt vesz a szexuális differenciálódás folyamatában, szoros kölcsönhatásban egy másik génnel, amelyet Z-génnek neveznek, amelynek funkciója általában a hím gének gének gátlásában áll. A normál 46XY hím genotípus esetében a SRY gén olyan proteint kódol, amely gátolja a Z gént, és specifikus hím gének aktiválódnak. A 46XX normál női genotípus esetében, amelyben a SRY hiányzik, a Z gén aktiválódik, és gátolja egy specifikus hím gént, ami feltételeket teremt egy női típus kialakulásához.
A nemi meghatározás kromoszóma elmélete
Fontolja meg a kromoszómális nemi meghatározást. Ismeretes, hogy a kétnemzetiségű szervezetekben (állatok és kétszínű növények) a nemek aránya általában 1: 1, azaz a férfiak és a nők egyaránt gyakori. Ez az arány egybeesik az elemző keresztben történő hasítással, amikor az egyik keresztezett forma heterozigóta (Aa), és a másik homozigóta a recesszív allélekre (Aa). Az utódok esetében ebben az esetben van egy hasítás az 1-hez képest Aa: 1aa. Ha egy nemet ugyanazon elv alapján örökölnek, akkor logikus lenne feltételezni, hogy egy nemnek homozigótanak kell lennie, a másiknak pedig heterozigótanak kell lennie. Ezután a nemek szerinti felosztásnak minden generációban meg kell egyeznie az 1,1-es értékkel, ami ténylegesen megfigyelhető.
A nemek evolúciós elmélete Az elmélet 1965-ben javasolt. Az elmélet egységes szempontból magyarázza a nemhez kapcsolódó számos jelenséget: szexuális dimorfizmust az egészségben és a betegségben, a nemek arányát, a differenciál mortalitást és a nemi válasz arányát, a nemi kromoszómák szerepét és a nemi hormonokat, agyi és kéz aszimmetriát, kölcsönös hatások, valamint a nemek közötti pszichológiai és társadalmi különbségek.
Az elmélet az aszinkron módon fejlődő konjugált alrendszerek elvén alapul. Férfi az operatív népesség alrendszer, női - konzervatív alrendszer. A környezetből származó új információk először elérik a férfi nemet, és csak sok generáció után kerülnek át a nőstényre, ezért a férfi nemi fejlődése megelőzi a nő fejlődését. Ez az időeltolás (két fázis a vonás evolúciója két vonásfajtát (hím és nő) teremt - a népesség szexuális dimorfizmusát. Az innovációk kereséséhez és ellenőrzéséhez szükséges az alrendszerek közötti evolúciós „távolság”.
 Hogyan jelenik meg az ember
Hogyan jelenik meg az ember Angol nyelvű írás egy barátról - új szókincs, példák és szöveg
Angol nyelvű írás egy barátról - új szókincs, példák és szöveg A legszokatlanabb természeti jelenségek
A legszokatlanabb természeti jelenségek