동물의 성 결정 유형. 성별 상속
인간의 성 결정 XY 메커니즘에 따라 발생합니다. 이 경우, 이종 성적인 섹스는 남성, 동성애 성적인 여성입니다. 성 결정은 염색체, 생식선 및 표현형의 세 단계로 나뉩니다.
포유류의 성별을 결정하기위한 두 가지 기본 규칙
고전적 배아 발생 연구는 포유류에서 성별을 결정하는 두 가지 규칙을 확립했다. 그들 중 첫 번째는 초기 토끼 배아에서 미래 생식선 (생식선 쿠션)의 세균을 제거하기위한 실험을 기반으로 1960 년대 Alfred Jost에 의해 공식화되었습니다. 생식선이 형성되기 전에 쿠션을 제거하면 여성으로 모든 배아가 발달했습니다.. 수컷 생식선은 태아 남성화를 담당하는 이펙터 호르몬 테스토스테론을 분비하고 그러한 해부학 적 변형을 직접 제어하는 \u200b\u200b제 2 안티-뮬러 호르몬 (MIS) 이펙터의 존재를 예측한다고 제안되었다. 관찰 결과는 원칙적으로 정립되었습니다 : 고환 또는 난소에서 생식선 발달의 전문화는 배아의 후속 성적 분화를 결정합니다.
1959 년까지, X 염색체의 수는 포유류에서 성 조절에 가장 중요한 요소 인 것으로 가정되었습니다. 그러나 단일 X 염색체를 가진 여성으로 암컷으로 발달 한 유기체와 남성으로 개발 된 하나의 Y 염색체와 다중 X 염색체를 가진 유기체의 발견은 그러한 아이디어를 포기해야했습니다. 포유류의 성별을 결정하는 두 번째 규칙은 다음과 같이 공식화되었습니다. Y 염색체는 남성의 성별을 결정하는 데 필요한 유전 정보를 가지고 있습니다..
위의 두 규칙의 조합을 때때로 성장 원칙이라고합니다. Y 염색체의 존재 또는 부재와 관련된 염색체 성은 배아 생식선의 분화를 결정하여 신체의 표현형 성을 조절합니다. 성별을 결정하는 비슷한 메커니즘을 유전이라고합니다. GSD) 환경 요인의 통제 역할에 근거한 것과 대조한다 (Eng. ESD) 또는 성 염색체와 오토 솜의 비율 CSD).
호르몬 성 결정
성 결정은 릴레이 경주의 형태로 표현 될 수 있으며, 염색체 메커니즘은 남성 또는 여성 생식기 기관으로 발달하는 미분화 생식선으로 전달됩니다. 생식선 발달에서 성 염색체의 역할을 연구 할 때 Y 염색체의 존재 또는 부재가 인간에게 결정적이라는 것이 밝혀졌습니다. Y 염색체가 없으면 생식선이 난소로 분화되어 여성이 발달합니다. Y 염색체가 있으면 수계가 발달합니다. 분명히, Y 염색체는 고환 분화를 자극하는 물질을 생성합니다. "자연의 기본 계획은 여성을 만드는 것이었고, Y 염색체를 추가하면 남성이 변하는 것으로 보인다." 계전기의 다음 단계는 태아의 성적 분화 과정과 해부학 적 발달을 결정하는 호르몬에 의해 계속됩니다. 출생시 프로그램의 첫 부분이 종료됩니다. 출생 후 계전기는 성의 형성을 완료하는 환경 적 요인으로갑니다. 일반적으로 항상 유전 적성에 따라 그런 것은 아닙니다. 성 결정은 복잡한 다단계 과정이며 생물학적뿐만 아니라 사람의 심리 사회적 요인에 달려 있습니다. 이것은 성전환, 이성애자, 양성애자 또는 동성애 행위 및 생활 방식으로 이어질 수 있습니다.
생식선 성 결정의 생리적 기초
성 결정 메커니즘의 생리적 기초는 포유류 배아 생식선의 양성입니다. 이러한 실행에서 Mueller 덕트와 Wolf 채널이 동시에 존재합니다-각각 생식 관의 기초는 암컷과 수컷입니다. 일차 성 결정은 Sertoli Cell이라는 달리기에 특수한 세포주가 나타나는 것으로 시작됩니다. 후자는 Jost에 의해 예측 된 안티-뮬레 리아 호르몬을 합성하는데, 이는 미래 난관의 자궁과 자궁 인 뮬러 덕트의 발달을 직접 또는 간접적으로 억제하는 역할을합니다.
유전자 성별 결정 메커니즘
SRY 유전자의 국소화를 갖는 인간 Y 염색체
1987 년 David Page와 그의 동료들은 280,000 개의 뉴클레오타이드 쌍의 길이를 가진 Y 염색체의 특정 단편과 염색체 사이의 부위 교환의 결과로이 영역을 포착하는 결실을 가진 암컷 XY를 물려받은 남성 XX를 탐색했습니다. 이 단편은 유 테리아의 모든 이들 동물의 Y- 염색체에 존재하고 유사 상 염색체 영역의 경계로부터 1 억 쌍의 뉴클레오티드 거리에 위치하며, ZFY 유전자의 길이는 1 억 쌍의 뉴클레오티드 쌍이다.
ZFY 상 동체-ZFX 유전자는 X 염색체에서 발견되며 ZFX는 비활성화되지 않습니다. ZFX 및 ZFY 인자는 모두 DNA- 결합 활성을 갖는 징크 핑거 모티프를 함유하는 전사 인자를 암호화한다. 성별 역전이있는 개인에서 Y 염색체의 특정 서열에 대한 추가의 상세한 분석은 검색을 35 kbp의 영역으로 제한했으며, \u200b\u200b고전 영어와 동등한 것으로 간주되는 유전자의 발견으로 이어졌습니다. 고환 결정 요인. 이러한 유전자를 SRY라고합니다. 성별 결정 영역 Y 유전자).
스리 성 결정 분야에 위치하며 80 개 아미노산 잔기의 단백질을 암호화하는 보존 적 도메인 (HMG 박스)을 함유한다. SRY 유전자의 활성은 생쥐에서 배아 발생 10-12 일에 고환으로의 분화 기간이 시작되기 전에 주목되었으며, 적어도이 단계에서는 생식 세포의 존재에 의존하지 않습니다. XY 여성에서이 유전자의 HMG 박스에서 특이점 돌연변이 또는 결실은 성별 역전을 초래한다. 마이크로 인젝션에 의해 측면 영역을 갖는이 유전자를 함유하는 14 kbp DNA 단편의 동종 동물 개체의 수정란으로의 이동은 핵형 XX를 갖는 수컷의 출현을 초래 하였다.
SRY 유전자 기능
SRY 유전자의 HMG 박스에 의해 암호화 된 도메인은 DNA에 특이 적으로 결합하여 구부러지게한다. SRY 단백질 또는 HMG 도메인을 함유하는 이의 동족체에 의해 야기 된 DNA 굽힘은 상당한 거리에 걸쳐 기계적으로 전달 될 수 있고 전사, 복제 및 재조합의 조절에 중요한 역할을한다. SRY가 국소화되는 DNA 영역에는 남성 유형에 의한 1 차 생식선 분화에 관여하는 주요 효소를 암호화하는 2 개의 유전자가 포함되어 있습니다 : P450 아로마 타제 유전자, 테스토스테론의 에스트라 디올로의 전환 및 밀러 덕트의 발달을 억제하는 요인으로 역전의 원인이되고 고환의 분화를 촉진하는 요인 .
또한 SRY 유전자의 산물은 Z 유전자라고 불리는 다른 유전자와 긴밀한 상호 작용으로 성적 분화 과정에 참여하며, 그 기능은 일반적으로 특정 남성 유전자를 억제하는 것입니다. 정상적인 남성 유전자형 46XY의 경우, SRY 유전자는 Z 유전자를 억제하는 단백질을 암호화하고 특정 남성 유전자가 활성화된다. SRY가없는 정상 여성 유전자형 46XX의 경우, Z 유전자가 활성화되고 특정 남성 유전자를 억제하여 여성 유형에 따라 발달 조건을 생성합니다.
성 결정의 염색체 이론
염색체 성 결정을 고려하십시오. 이성 유기체 (동물 및 이성 식물)에서 성별 비율은 일반적으로 1 : 1, 즉 남성과 여성 개인이 똑같이 자주 발견되는 것으로 알려져 있습니다. 이 비율은 교차 형태 중 하나가 이형 접합 일 때 분석 교차에서의 절단과 일치합니다. (Aa) 다른 하나는 열성 대립 유전자에 대한 동형 접합체입니다 (aa). 자손의 경우이 경우 관련 분할이 관찰됩니다. Aa : 1aa. 성별이 동일한 원칙에 의해 상속된다면, 한 성별이 동형 접합이고 다른 이형 접합이어야한다고 가정하는 것이 합리적입니다. 그런 다음 성별 분할은 각 세대에서 1.1과 같아야하며 이는 실제로 관찰됩니다.
진화론 적 성 이론 이론은 V. Geodakyan에 의해 1965 년에 제안되었다.이 이론은 성과 관련된 많은 현상들 : 규범과 병리학에서의 성적 이형성, 성비, 차별 사망률 및 성 반응의 규범, 성 염색체와 성 호르몬의 역할, 뇌와 손의 비대칭 성, 상호 효과, 남녀 간의 심리적, 사회적 차이.
이론은 비동기 적으로 진화하는 공액 서브 시스템의 원리를 기반으로합니다. 남성 성별은 운영 인구의 하위 시스템, 여성 보수적 인 서브 시스템. 환경으로부터의 새로운 정보는 먼저 남성 성별로 가고, 많은 세대가 여성에게 전달 된 후에야 남성 진화가 여성 진화에 선행한다. 이번 타임 시프트 (2 개 단계 특성의 진화)는 두 가지 형태의 특성 (남성과 여성)을 만듭니다-인구의 성적 이형성. 혁신을 찾고 검증하기 위해서는 서브 시스템 사이의 진화적인“거리”가 필요합니다.
성별- 그것은 생식을 결정하는 형태 학적, 생리 학적, 생화학 적, 행동 및 다른 징후의 조합이다.
성별이 다른 개인의 징후는 일차 및 이차로 나뉩니다. 일차는 gamete 형성과 수정을 제공하는 기관 (gonads, 생식기 기관)으로 표현됩니다. 이들은 외부 및 내부 생식기이며 배 발생에 놓여 있습니다. 보조-성적 복제에 참여하지 마십시오. 성 호르몬의 영향으로 발달하고 사춘기 동안 (12-15 세의 인간에서) 나타납니다. 이것들은 근골격계 발달, 피하 지방, 모발, 음성 음색, 행동의 특징입니다. 새에서-노래, 깃털 등
성별과 관련된 개인의 특성은 3 가지 범주로 나눌 수 있습니다.
1) 바닥으로 제한
2) 성 통제
3) 바닥에 결합.
전자의 발달은 두 성별의 상 염색체에 위치한 유전자에 기인하지만 한 성별에서만 발생합니다. 따라서 알을 낳는 유전자는 닭과 수탉에 있지만 닭에만 나타납니다. 소의 우유와 우유 유전자도 비슷하게 행동합니다. 이 현상은 해당 성 호르몬에 대한 노출과 관련하여 관찰됩니다.
두 번째 예는 수컷의 암소에서 뿔이 나타나고 여성에서는 거룩함이 나타납니다. 인간의 경우 : 남성의 대머리, 통풍-남성의 80 %, 여성의 12 %.
성 염색체의 유전자에 의해 발달이 조절되는 특성을 성관계라고합니다. 색맹, 혈우병은 X 염색체와 관련이 있습니다. Y- 염색체-고혈압증, ichthyosis.
층의 결정 유형.
1. 대충-수정 될 때까지. 성 염색체의 비율은 이것에 영향을 미치지 않습니다. 난 모세포는 이배체이다. (일부 웜, 로티퍼-암컷은 큰 난 모세포에서, 수컷은 작은 난 모세포에서 발생합니다).
2. Singamny-수정 중 성의 유전자 결정. 성 염색체의 조합 특성 또는 성 염색체와 오토 솜의 비율에 따라 다릅니다.
3. Epigamous-외부 환경 (bonelia worm)의 영향을받습니다.
층의 염색체 결정
┌─────────────────────────┬───────────────────┬──────────────────┐
│ 염색체 종류 │ 유전자형 │ gametes 종류 │
│ 성별 정의 ├─────────┬─────────────────────────────
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
│ 남성 이질성 │ │
│ Orthoptera 곤충 │ │ │ │ │
│ (프로 테너 버그, 버그, │ │ │ │
│ 거미, 메뚜기) │ ХО │ ХХ │ Х, О │ Х │
│ 초파리 │ XY │ XX │ X, Y │ X │
│ 무척추 동물 │ │ │ │ │
│ (포유류, 인간) │ XY │ XX │ X, Y │ X │
│ │ │ │ │ │
│ 여성 이질성 │ │
│ 새. 물고기, 나비, │ │ │ │ │
│ 누에, 파충류, 젬 -│ │ │ │ │
│ 물. │ XX │ XY │ X │ X, Y │
│ 나방 및 기타 거꾸로 │ │ │ │ │
야간 │ XX │ XO │ X │ X, O │
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
유전자 제어를 사용한 염색체 성 결정은 동종 이형 결정에 속합니다. 섹스를 담당하는 사람들을 섹스 염색체라고합니다. 정상적인 수컷 생식 세포는 X 또는 Y 염색체를 운반하고 모든 알은 X 염색체를 운반합니다. 감수 분열 중 정상적인 염색체 발산의 경우, 일반적인 염색체 X와 Y의 세트를 갖는 정상적인 난자와 정자 세포가 형성되며, 접합자 성별은 균주 XX와 XY의 비율에 의해 결정됩니다 (동종 및 이종).
성의 염색체 이론 (1907)은 수정 중 성 염색체의 조합에 의해 성별이 결정된다는 것입니다. 다음 유형의 염색체 성 결정이 구별됩니다 : XY, XO, ZW, ZO.
유사 분열 또는 감수 분열의 과정이 방해되면 gynandomorph 개인이 형성 될 수 있습니다. 이러한 개체의 다른 세포에서 성 염색체의 함량은 다를 수 있습니다 (모자이크). 초파리 파리에서 : XX와 XO, 인간의 경우 XX와 XY, 신체의 다른 부분이 대응하는 성 특성을 가질 수 있습니다. 다른 모자이크 현상이있을 수 있습니다 : XX / XXX, XY / XXX; XO / XXY 등
성 염색체가 갈라지지 않으면, 인간 염색체에 성 염색체의 12 가지 가능한 조합이있을 수 있으며, 이것이 인간 염색체 이상의 원인입니다.
│ X │ XX │ 오
─────┼────────┼─────────┼────────
X │ XX │ XXX │ XO
Y │ XY │ XXY │ 요
XY │ XXY │ XXXY │ XYO
O │ XO │ │
감수 분열 동안 성 염색체가 발산되지 않는 경우, 암컷에서 생식선 XX 및 O가 형성된다. 남성의 XY 및 O. 수정에 참여하면 성 염색체의 특이한 조합으로 접합자가 형성됩니다. 인간의 경우 이러한 이상은 600-700 명의 신생아 중 1 명에서 발생합니다. Zygote YO는 초기 단계에서 사망합니다. 개인 ХХХ, ХХY, ХО는 실행 가능하며 성별은 "Y"염색체의 존재 유무에 따라 달라집니다.이 염색체는 X 염색체의 수에 따라 남성의 성 형질의 형성, 발달 및 고환의 형성을 조절합니다. 과도한 X 염색체는 체질 이상 및 지적 결함을 유발합니다. 그러나 본질적으로 "Y"염색체가 유 전적으로 불활성이며 성 결정에 특별한 영향을 미치지 않는 개체가 있습니다.
초파리에서 수컷이지만 불임 인 (1916 년, K. Bridges) XO 유형의 개체가 발견되었으며, XXY 개체는 정상적인 다산 암컷이었다.
균형 잡힌 성 이론 (Bridges, 1922). 생식기와 오토 솜의 비율이 연구되었습니다.
2n 염색체 세트가있는 정상 암컷에서, 오토 솜과 X 염색체의 비율은 1 : 2n \u003d 2A + 2X (2X : 2A \u003d 1은 정상 암컷), 암컷에 비해 1, 5 : 2A + 3X (3X : 2A \u003d 1, 5) 비옥 한). 수컷의 경우 비율은 0.5 2n \u003d 2A + XY (X : 2A \u003d 0, 5)입니다. 개인의 감소와 함께 남성 3A + XY (X : 3A \u003d 0, 33-불모의) 수퍼 남성으로 남아 있습니다. 1과 0, 5 사이의 계수는 _intersex. : 3A + 2X (2X : 3A \u003d 0, 66-두 성별의 증상, 무균)에서 중간체의 표현형에 해당합니다.
따라서 균형 이론의 본질은 성 염색체뿐만 아니라 오토 솜이 성 결정에 참여한다는 것입니다. 하나의 반수체 세트의 오토 좀이 수컷을보고합니다. 이 경우 섹스는 오토 좀 및 성 염색체의 수 (균형)의 비율에 의해 결정됩니다.
배수성에 의한 성 결정은 꿀벌에서도 발견됩니다. 여성 꿀벌은 이배체이고 수컷은 반수체입니다. 수정되지 않은 난에서 부분 생식으로 발달합니다.
종양 발생에서 층의 구별
초기 배아의 생식선 초반 (최대 5 주 또는 6 주까지)은 다른 성별 사이에 다르지 않으며 외층-피질 피질 및 내층-수질로 구성되며, 생식 세포를 포함하지 않습니다. 배아 경로의 일차 세포는 노른자 낭의 외배엽에서 배아 발생 3 주째에 인간에서 발견된다. 그런 다음 화학 신호의 영향으로 생식선으로 이동합니다. 이 이주는 성별에 독립적입니다. 생식선 초본은 난소 나 고환에서 생길 수 있습니다. 생식선 분화는 8 주째에 관찰됩니다 : 36 일에 고환은 남성의 발달을 결정하는 안드로겐 (테스토스테론)을 분비하기 시작합니다.
생식선이있는 유기체에서는 성적 특성의 형성에 대한 유전자 제어가 수행됩니다.
성 분화는 암컷 및 수컷 생식 세포의 융합 동안 형성된 성 염색체의 유전자 세트에 의해 프로그램된다. 배아의 유전자 성별은 성 염색체 XX 또는 XY에 의해 결정됩니다.
성 발달 방향은 Y 염색체의 존재에 의해 결정됩니다. 일반적으로 X 염색체에는 남성 유전자의 발달을 막는 억제 유전자 (Tfm testicular feminization gene)가 포함되어 있습니다. 정상적인 유전자 대립 유전자는 남녀 모두에서 합성 된 안드로겐에 대한 단백질 수용체의 합성을 결정합니다. 수컷 표현형에 따른 발달은 Y- 염색체 유전자, H-y 항원에 의존한다 (1955 년 마우스에서 설명 됨; HA 유전자). 1 차 수컷 생식 세포 세포가이를 분비합니다. Y 5NA 0은 테스토스테론 (남성 성 호르몬)의 생성을 담당합니다. 이 세포가 생식선의 기초에 들어가 자마자 고환의 분화가 시작됩니다. H-Y 수용체는 두 가지 유형의 생식선 세포 표면에 존재합니다 (소의 성 발달과의 편차). 남성 표현형은 전체 남성 염색체에 의해 결정되는 것으로 여겨졌다. 그러나 1990 년에 Y 염색체의 핵형에 국한된 유전자 (Sex Region Y)가 발견되었습니다. 부재시, XY 유전자형은 여성 표현형을 제공한다.
고 나드 개발
│ sexual │ 배아 양성의 생식선
│ ├┼┼┼┤ │ 알
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
유전자형이 XX │ │ 유전자형이 XX 인 경우
(7-8 주) ┌──┴──┐ ┌───┴───┐ (6 주차)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
피질 │ └┴┘ │ │└┴┴┴┴┴┘│ 수질 층
레이어 └─────┘ └───────┘
(유전자는 여성의 성별을 결정) (유전자는 남성의 성별을 결정)
사람의 기능을 조절하는 특정 여성 호르몬이 있습니다.
표시가 눈에 띄게되고 삽입 광고의 산물
우리는 7-8주의 끝에. 난소 또는 정액 세포의 20 번째 조직
(Leydig 세포)를 포함한 1 차 생식 세포. 그들은 형성
호르몬 성 (테스토스테론,
생식선 및 에스트라 디올과 차별화). 10-12 주에
두 번째 달 말까지 난소. 내부 성기
우리의 깊이에서 3 개월 말까지, 12 주에 특정
배아 생식선은 배아의 안드로겐 수준에서 감지됩니다
난 모세포 (prophase MI). 차이-남성화 시작
중간 층의 7 개월까지 난소 순환) 및 완료-
20 주째 난소에서 9 개월 째에 녹습니다.
200-400 천개의 난 모세포 II가있다. 사춘기에서는 est의 수준
로겐은 증가하고 영향을 미칩니다
골격의 구조와
안드로겐 (각각
여성 및 남성 유형).
생식선은 1 차적인 성적 특성과 2 차적인 발달을 결정합니다. 성선은 내분비선의 호르몬과 함께 성 분화를 조절하는 호르몬 (에스트라 디올, 안드로겐)을 분비합니다. 호르몬 수준은 차례로 유전자에 의해 제어됩니다.
따라서 성적 분화 과정에는 다음이 포함됩니다.
1) 유전자 제어;
2) 호르몬의 조절 기능.
유전자에 대한 조절 인자로서 호르몬의 작용 이론이 있습니다. 그들은 특정 표적 세포에서만 작용합니다. 특별한 단백질이 세포에서 생성되는데, 호르몬에 결합하고 동시에 변화하는 수용체는 염색체에서 하나 또는 여러 유전자의 작용을 유도하는 특성을 얻습니다. 자궁 세포에 대한 글루코 코르티코이드의 영향으로 RNA와 단백질의 합성이 변화합니다 (구성표 참조).
수용체 단백질 및 호르몬의 형성은 유전자에 의해 제어된다. 통제를 위반하는 경우 이상이 발생할 수 있으며 그 예로 모리스 증후군이 있습니다. 고환 여성화 (Morris syndrome) (Morris, 1953) :이 질환을 가진 개인은 테스토스테론 수용체가 부족합니다. 안드로겐은 정상적인 양으로 분비됩니다. 모리스 증후군에서 배아 발생은 남성 성 호르몬을 생성하기 시작하는 고환을 동반합니다. 그러나 이러한 배아에서는 수용체 단백질 (열성 유전자 돌연변이)이 형성되지 않아 남성 성 호르몬에 대한 장기 발달 세포의 감도가 보장됩니다. 이로 인해 남성 유형의 발달이 멈추고 여성 표현형이 나타납니다. 예외적 인 경우 적절한 호르몬을 도입하여 그러한 결함을 교정 할 수 있습니다.
따라서 원시 유전자 양성은 성별 재정의의 기초입니다. 남성 태아는 여성 성별의 특징적인 특성을 얻습니다. 남성 핵형, 남성 생식선, 여성 표현형. 신체 비율은 여성이며 유선, 유선 질 단축, 음순의 고환, 사타구니 운하 및 복강이 있습니다.
층 재정의
호르몬에 노출되거나 표적 세포 수용체의 병리로 인해 성별 재정의가 발생할 수 있습니다 (Moris 증후군, 수탉의 성선 제거).
실제로 성의 발달을 통제하는 유전자의 효과를 약화시키는 많은 요소가 있습니다. 예를 들어, 한 개인의 생식선에있는 사람은 고환과 난소 부분을 고르게 발달시킬 수 있습니다 (고환과 난소가 발달합니다) hermaphroditism은 성애의 현상입니다.
임상 데이터를 기반으로 세 가지 유형의 성교가 구별됩니다.
1) 진정한 hermaphroditism : 남녀의 생식 세포의 존재;
2) 수컷 유사성 간염 : 고환 (고환, 고환), 여성 표현형 만 있습니다.
3) 여성 pseudohermaphroditism : 난소, 남성 표현형이 있습니다.
이 분류는 세포 유전 학적 기초와 일치하지 않기 때문에 한 사람의 46 XY에는 옵션이 있습니다.
성비.
기본 성비 (성 염색체의 만남도 마찬가지로 가능하므로 수정시 1 : 1에 가까워 야합니다). 인간을 대상으로 한 조사에 따르면 100 개의 여성 접합자마다 140-160 개의 남성 접합자가 형성되는 것으로 나타났습니다. Y 염색체를 함유 한 정자는 더 가볍고, 더 이동성이 있으며, 큰 음전하 (알은 양전하를))가 있습니다. 따라서 Y 함유 정자는 종종 난자를 수정합니다.
2 차-출생시까지 103-105 명의 소년이 100 명의 소녀마다 태어납니다. 여성 생식 세포의 생존력, 남성 배아 단백질의 이물질. 20 세가되면 100 명의 소녀 당 100 명의 소년이 있습니다.
고등 교육 – 50 세까지는 100 명의 여성 당 85 명의 남성이 있으며, 85 세까지는 100 명의 여성 당 50 명의 남성이 있습니다. 여성의 몸은 성 염색체에 의한 여성의 모자이크로 다른 이유와 함께 설명 될 수 있습니다.
성 염색체에 대한 여성 모자이크에 대한 가설 M. Loyon.
1949 년에 M. Borr와 C. Bertrand는 여성 신경 세포의 핵에서 염색 된 염색질 블록이 발견되었다는 것을 발견했습니다. 사람 세포의 핵에서는 그것을 감지하지 못합니다. 이 덩어리를 성 크로 마틴 (Borr 's body)이라고하며 비활성화 된 X 염색체를 나타냅니다.
발달 초기에 X 태아 염색체는 암컷 태아에서 작용합니다. 사람의 유전자보다 두 개 더 이것은 암컷 접합자의 큰 생존력을 설명 할 수 있습니다.
1962 년 M. Lyon은 암컷 포유 동물 유기체에서 하나의 X 염색체의 불 활성화를 가정했습니다. 암컷 태아에서 두 염색체는 최대 16 일의 배아 발달 기능을합니다. 16 일째에 성 염색질의 형성으로 하나의 염색체가 비활성화됩니다. 이 과정은 임의적이므로, 약 1/2 세포에서 모체 X 염색체 X 5M 0은 활성 상태를 유지하고 부계는 비활성화됩니다. 다른 경우에는 아버지가 활성화되고 (X 5 O 0) 어머니가 비활성화됩니다. 재 활성화가 발생하지 않습니다. 모계 및 부계 X 염색체는 대립 유전자이지만 절대적으로 동일한 유전자는 포함하지 않습니다. 우성 대립 유전자는 하나의 염색체에 위치하고 열 성체는 다른 염색체에 있습니다. 추가 유전자를 소유하면 신체의 적응 능력이 확장됩니다.
성적 차이 성적 차이 진단
차별화
┌─┐ ┌┴┐
게임 X X └┬┘ └─┘ Y
난자 --┼─ 정자
┌───────────────────┐
유전자 │ 염색체 XX 및 XY │
└───────────────────┘
┌─────────┴─────────┐
생식선 │ 미분화-│
┌── │ 태아 생식선 │
│ └───────────────────┘
│ │ HY 항원
│ ┌────────┴────────┐ ┌─────────────┐
생식기 │ │ │ │1 생식기 │
숙성 │ │ │ ┌─│ 표시 │
│ │ │ │ └────────────┘
│ ┌─────┴─────┐ ┌────────────┐ │ u │
│ │ 고 나드 │ │ 고 나드 ├────┤ 7-32 주
│ │ 난소 │ ен 고환 │ анд 안드로 호르몬
│ │ └───────────┘ └────────────┘ │ 세포 유전자
│ │ │ 선도
│ │ 사춘기 │ │ ┌─────────────┐
│ └───────────────────────────────┘ │ │ 미분 │
호르몬 │ 뇌하수체 호르몬, 남녀 │ │ 신경-│
│ 호르몬 : 안드로겐 (테스토스테론), 에스트로 -└─│ 새로운 방법 │
└─ 유전자 (프로게스테론) │ 성별 │
┌──────────────┴───────────────┐ └────────────┘
표현형 │ 이차 성적인 특성 │
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
심리적 │ 성별과 행동 │
└───────────────────────────────────────────────────┘
여성의 몸은 추위, 전리 방사선, 정서적 과부하에 더 강합니다 (여성은 더 자주 울고 활성 아민은 눈물과 함께 방출되어 결과적으로 혈압이 감소합니다).
가설이 제한없이 작동한다면, 2 개의 X 염색체를 가진 건강한 여성과 X 4 0을 가진 환자 또는 XY / XXYY의 남성 사이에 표현형 적 차이가 없을 것입니다. 분명히, X 염색체는 완전히 비활성화되지 않습니다.
성별 비율의 규제.
정자가 일정한 전기장에 놓이면 X와 Y 염색체가 분리됩니다. 축산에 사용됩니다. 올바른 성의 후손의 80 %를 얻는 것이 가능합니다.
남성 또는 여성의 성별에 대한 인식은 성별, 정신적 매개 변수에 대한 정신적 인식의 필수 부분입니다. 성전환-심리적 유두 염, 사람의 성적 정체성을 위반하는 현상. 이성에 속한다는 사람들의 강박 적 인식 사례는 고대부터 알려져 왔습니다. 그래서 헤로도투스는 그의 미스터리 인“사이 티병 (Scythian disease)”에서 여성복을 입었을뿐만 아니라 이성의 성격 적 특성을 채택한 남자를 묘사했다. 일부 황제들은 옷을 입는 데 어려움을 겪었습니다 (예 : Caligula). J. D”호.
해부학 적 및 정신적 매개 변수의 불일치는 내부 환경과 사회의 충돌입니다. 여성의 영혼은 남성의 몸에 살고 그 반대도 마찬가지입니다. 외과 성 교정이 이루어집니다. 남자를 여자로 바꾸려면 한 번의 수술이 필요합니다. 성기와 음낭의 피부에서 질이 형성됩니다. 3-4 작업 (유방 땀샘 제거, 음경 형성) : 여자에게서 남자를 만드는 것이 더 어렵습니다. 이 사람들은 영원한 고통에 처해질 것입니다 : 그들은 호르몬 약을 마시고 아이들이 없습니다. 모스크바에서 인간 복제 및 가족 계획 연구소.
성적 분화에는 일차 (고 나드)와 이차 성적인 특성이 형성됩니다.
대부분의 동물에서 성별은 수정시 유전자 적으로 결정됩니다.
X 염색체는 고환 여성화 유전자 (Tfm), 정상을 가지고 있습니다
안드로겐에 대한 단백질 수용체의 합성을 결정하는 대립 유전자,
암컷과 수컷 유기체에서 합성됩니다.
┌──────┐-각 염색체는 non-senegen Tfm입니다-
치트가 피질을 개발 중입니다.
Y────┤ └┴┴┘ ├──────┐ XY-I 유전자 5NA-gene 0을 포함합니다.
│ └──────┘ │ 항원 합성 정의
│ M │ 수컷 \u003d 생산 책임
│ │ 테스토스테론.
┌──┴──┐ ┌───┴───┐ 생식선 분화는 7-10에서 관찰됩니다
│┌┬┬┬┐│ │ │ 일주일. 10 주차에는 바닥이 운영 될 수 있습니다
│└┴┴┴┘│ │ │ │ 염색체 세트별로 정렬합니다.
└─────┘ └───────┘
발달 방향은 Y 염색체의 존재에 의해 결정됩니다. 일반적으로 XX 염색체에는 남성 유전자의 발달을 막는 억제 유전자가 포함되어 있습니다. 이 발달은 N-Y (HA) 항원에 달려 있으며, I 5HA 유전자는 테스토스테론 생산을 담당합니다. 여러 종에서 발견되었다 (1955 년 마우스, Silcher의 Eichwald). 1 차 수컷 생식 세포 세포가이를 분비합니다.
일반적으로 성 염색체의 조합을 가진 개인에서 여성 유형은 여성 성별을 결정하는 유전자와 남성-남성 성별에 의해 지배됩니다.
생식기는 1 차 신장에서 나오는 Mullerian과 Wolf 덕트에서 형성됩니다. 여성의 경우 ül 러 덕트는 나팔관과 자궁으로 발달하는 반면 늑대 덕트는 위축됩니다. 남성의 경우 늑대 덕트는 정관과 정낭으로 발달합니다. 배아 고환에서 스테로이드 호르몬 인 테스토스테론과 5- 디 하이드로 테스토스테론은 모 호르몬 (코로 이온 성선 자극 호르몬)의 영향으로 합성됩니다. 이 호르몬은 생체 전위에 작용합니다
외부 및 내부 생식기 : Wolf ducts, Mullerian ducts 및 urogenital sinus. 이러한 요소가 모두 작동하면 정상적인 남성의 신체가 발달합니다. 부재시 여성의 성적 특성이 형성됩니다. 남성과 남성 표현형의 불완전한 개발
유전자형 (남성 슈도 허 식염).
현대 생식 전략 :
-인공 수정;
-시험관 내 수정;
-배아의 인공 배양 및 자궁으로의 이식;
-대리모.
대부분의 동물은 부패한 유기체입니다. 성은 자손의 번식과 유전 정보의 전달 방법을 제공하는 특성과 구조의 조합으로 간주 될 수 있습니다. 섹스는 수정시 가장 자주 결정됩니다. 즉, 접합 핵형은 섹스를 결정하는 데 중요한 역할을합니다. 각 유기체의 핵형에는 남녀 (남성)가 서로 다른 성 염색체 (성 염색체)와 염색체 (여성)가 동일한 염색체가 포함되어 있습니다. 인간에서, "여성"성 염색체는 2 개의 X 염색체입니다. 생식 세포가 형성 될 때, 각각의 난은 X 염색체 중 하나를 받는다. X 염색체를 갖는 동일한 유형의 생식자가 형성되는 성별을 동종 게임이라고합니다. 인간에서 여성의 성별은 동성애자입니다. 인간의 "남성"성 염색체는 X 염색체 및 Y 염색체입니다. 생식 세포가 형성 될 때 정자의 절반은 X 염색체를 받고 다른 절반은 Y 염색체를받습니다. 다른 유형의 생식자가 형성되는 섹스를 이종 게임이라고합니다. 인간에서 남성의 성별은 이질적입니다. 두 개의 X- 염색체를 운반하는 접합체가 형성되면, X- 염색체 및 Y- 염색체가 수컷이면 암컷 유기체가 형성됩니다.
동물에서 다음과 같은 4 가지 유형의 염색체 성 결정.
1. 여성의 성별은 동성애 (XX)이고, 남성은 이종성 (XY) (포유류, 특히 인간, 초파리)입니다.
인간의 염색체 성 결정의 유전 체계 :
Drosophila의 염색체 성 결정의 유전 체계 :
2. 여성의 성별은 동성애 (XX)이고, 남성은 이종 (X0) (인간)입니다.
사막 메뚜기에서 염색체 성 결정의 유전 체계 :
3. 여성의 성은 이질적 게임 (ХY)이고, 수컷은 동성애 게임 (XX) (새, 파충류)입니다.
비둘기의 염색체 성 결정의 유전 체계 :
Yandex.Direct 모든 광고
4. 여성의 성별은 이질적 (X0)이고, 수컷은 동성애 (XX) (일부 곤충)입니다.
동물의 대다수는 남성과 여성의 두 성별의 개인으로 표현됩니다. 일부 식물 종에서는 두 성별의 개체도 관찰됩니다 (대마, 간 구상 동맥 이끼, 졸음 등). 성의 유전 법칙에 대한 일부 관찰이 훨씬 일찍 이루어 졌음에도 불구하고 유전 염색체 이론을 만든 후에 개인의 성별을 결정하는 메커니즘이 분명해졌습니다. 종의 개체군에서 평균적으로 남성과 여성의 비율은 동일합니다. 즉 1 : 1과 같습니다. G. 멘델에서의 이러한 비율은 상기 한 바와 같이 모노 하이브리드 분석 크로스와 유사성을 일으켰다. (Aa)다른 하나는 열성에 대한 동형 접합체입니다 (aa).성별로 분할 할 때 관찰 된 1 : 1 비율은 성별 중 하나가 이형 접합성이고 다른 하나는 신체의 성별을 결정하는 요인에 대해 동형 접합성이라고 가정 할 수있었습니다. 세기 초의 이러한 가정은 유전자 실험의 결과에 의해 확인되었고 성 염색체의 발견 후 세포 학적 확인을 받았다. 동물의 생식 세포와 신체 세포의 염색체 구조를 연구 할 때, 수컷과 암컷은 염색체 세트가 다르다는 것이 밝혀졌습니다. 성 염색체는 W. Wilson과 C. Mack Klang에 의해 자세히 연구되었습니다.
성 염색체. Drosophila 염색체의 성별을 결정하기위한 염색체 메커니즘의 현재 아이디어를 고려하는 것이 편리합니다. 그림. 도 84는 4 쌍을 포함하는 8 개의 염색체로만 이루어진 초파리 염색체 세트의 개략도이다. 세 쌍으로, 염색체는 서로 상 동성이며 형태 학적으로 다르지 않습니다. 네 번째 쌍의 염색체는 매우 다릅니다. 그 중 하나 인 X- 염색체는 막대 모양이고, 두 번째 (Y- 염색체)는이 염색체의 불균등 한 팔에 의해 형성된 고리 모양입니다 암컷 세포에서는 초파리의 네 번째 쌍이 두 개의 X- 염색체로 나타납니다

하나 X 하나의 Y 염색체. 수컷과 암컷을 구별하는 한 쌍의 염색체를 성 염색체 또는 이종 염색체라고하며, 다른 염색체는 오토 좀이라고도합니다 (때로는 A로 약칭 함).
동성애와 이성애 섹스. 같은 성 염색체를 가진 초파리 여성에서XX), 감수 분열로 인한 모든 생식 세포는 동일한 성 염색체 (X)의 결과로 모든 gamete가 동일합니다. Drosophila의 여성 섹스는 단 한 가지 유형의 gametes를 생산하는 것을 homogametic이라고합니다. Drosophila 수컷은 두 가지 종류의 생식자를 생산하는데, 그중 하나는 성 염색체가 X 염색체로, 다른 하나는 Y 염색체로 표시됩니다. Drosophila의 남성 섹스는 따라서 이질적입니다.
동성 또는 이종성에 대한 암컷의 소속에 따라 모든 동물은 두 그룹으로 나뉩니다. 포유류, 벌레, 갑각류, 대부분의 곤충 (Drosophila 포함), 대부분의 양서류 및 일부 물고기에서 남성의 성별은 이질적이며 여성은 동성애 적입니다.
이 그룹에는 사람들이 포함됩니다. 인간 체세포에는 44 개의 상 염색체 염색체가 포함되어 있으며, 여성에게는 2 개의 X 염색체가 있고 남성에는 1 개의 X 염색체가 있으며 하나의 Y 염색체. 여성 인체의 염색체 공식은 44A +로 지정할 수 있습니다 XX, 수컷-44A + XU. 조류, 파충류, 일부 양서류 및 물고기에서 일부 곤충 (나비 및 caddis 파리)은 암컷입니다. 이 경우 성 염색체를 지정하는 데 다른 기호가 사용됩니다. 여성 이종 성으로 성 염색체는 Zw, 남성 성 염색체,- Zz. 예를 들어 체세포에 78 개의 염색체가있는 닭에서 수컷 염색체 공식은 -76A + Zz, 여성-76A + Zw.
일부 종에서는 진화 과정에서 이종 성의 성 염색체 중 하나가 손실되어 염색체 세트에 성 염색체가 하나만 존재합니다. 이종 성에서의 성 염색체 공식은 다음과 같이 표시됩니다. XO또는조.
바닥에서 분리하는 메커니즘 1 : 1. 성별을 결정하는 염색체 메커니즘에 대한 지식을 통해 남성과 여성 개인의 출현 원인을 1 : 1 비율로 설명 할 수 있습니다. 위에서 언급 한 바와 같이, 동성애 성은 같은 유형의 생식자를 형성하며, 각각은 동일한 성 염색체를 가지고 있습니다. 차례로, 이종 성의 게임에서 하나 또는 다른 성 염색체가 나타납니다. 예를 들어, 초파리에서 암컷의 모든 생식 세포에는 하나의 X 염색체가 있고, 수컷에있는 생식 세포의 절반에는 X 염색체가 있고, 나머지 반에는 Y 염색체가 있습니다.
계란이X)는 X 염색체로 정자에 의해 수정되며, 접합체에서 두 개의 X 염색체의 조합은 암컷을 형성합니다 (XX) U 염색체가있는 정자와 같은 난자를 수정하면 수컷 (XU)이 나타납니다. 하나 또는 다른 정자의 수정에 참여하는 것도 똑같이 가능하기 때문에 남성과 여성이 나타날 확률은 동일합니다. 다른 종의 동물에서 다수의 신생아 개체에서 성별을 설명하면 1 : 1 또는 매우 가까운 비율을 나타냅니다. 따라서 신생아 100 명 중 남성은 인간 51 명, 개 51 마리, 닭 49 마리, 생쥐 50 마리, 양 49 마리, 소 51 마리, 말 52 마리입니다. 출생시 다른 성별의. 앞으로 이러한 비율은 성별이 다른 개인의 고르지 않은 생존으로 인해 크게 달라질 수 있습니다. 따라서 인간의 경우 50 세까지 남성과 여성의 비율은 85 : 100, 85 세-50 : 100입니다.
닭, 누에 등에서 볼 수 있듯이 염색체 메커니즘에 의해 발생하는 성 (性)의 발현은 실험적으로 변화 될 수있다. 수컷과 암컷 식물을 가진 이성 식물에서는 성 염색체도 연구되어왔다. 이 식물들 대부분의 이질은 남성 성별입니다.
성별의 유전학
성별은 염색체에 위치한 유전자에 의해 결정된 복잡한 특성을 특징으로합니다. 인체 세포에서 염색체는 쌍성 이배체 세트입니다. dioecious 개인을 가진 종에서, 남성과 여성의 염색체 복합체는 동일하지 않으며 한 쌍의 염색체 (성 염색체)에 따라 다릅니다. 이 쌍의 동일한 염색체는 X (X) 염색체라고 불렀으며, 다른성에는 없었습니다-U (게임) 염색체; 차이가없는 나머지는 오토 솜 (A)입니다.
여성의 세포에는 두 개의 동일한 성 염색체가 있으며 XX로 지정되며 남성에서는 두 개의 짝이없는 염색체 X와 Y로 표시됩니다. 따라서 남성과 여성의 염색체 세트는 하나의 염색체 만 다릅니다. XY
인간에서 생식 세포의 분열과 성숙 동안, 반수체의 염색체를 가진 생식자가 형성됩니다. 알은 일반적으로 22 + X 염색체를 포함합니다. 따라서 여성에게는 한 가지 유형의 생식자가 형성됩니다 (X 염색체가있는 생식선). 남성의 경우, 생식 세포는 22 + X 또는 22 + Y 염색체를 포함하며 두 가지 유형의 생식 세포가 형성됩니다 (X 염색체가있는 생식 세포와 Y 염색체가있는 생식 체). 수정 중에 X 염색체가있는 정자가 난자에 들어가면 암컷 태아가 형성되고 수컷에게 Y 염색체가 형성됩니다.
따라서 사람의 성별 결정은 정자, 수정란, X 또는 Y 염색체와 같은 남성 생식 세포의 존재에 달려 있습니다.
염색체 성 결정에는 네 가지 주요 유형이 있습니다.
1. 남성의 성은 이기종입니다. gametes의 50 %는 X-, 50 % -U-염색체를 가지고 있습니다 (예 : 인간, 포유류, dipterans, 딱정벌레, 벌레) (슬라이드 4).
2. 남성의 성은 이질적이다. gametes의 50 %가 X-를 운반하고 50 %는 성 염색체가 없습니다 (예 : 메뚜기, 캥거루) (슬라이드 7).
3. 여성의 성은 이질적이다. gametes의 50 %는 X-, 50 % gametes – Y– 염색체 (예 : 조류, 파충류, 꼬리가있는 양서류, 누에)를 가지고 있습니다 (슬라이드 7).
4. 여성의 성은 이질적이다. gametes의 50 %는 X-를 가지고, 50 %는 성 염색체, 예를 들어 두더지가 없습니다.
유전자가 성 염색체에 국한된 형질의 상속을 상속이라고합니다. 바닥과 연동됩니다.
26. 통합 시스템으로서의 유전자형. 유전자의 상호 작용, 유전자의 다중 효과.
전체 론적 시스템으로서의 유전자형
유전자의 특성. 에 기초 데이트 모노 및 디 하이브리드 크로스에서 캐릭터의 상속 사례를 통해 유기체의 유전자형이 개별적으로 독립적으로 작용하는 유전자의 합계로 구성되어 있으며 각각의 특성이나 특성의 발달 만 결정한다는 인상을받을 수 있습니다. 형질과 유전자를 직접적이고 명확하게 연결한다는 아이디어는 현실과 일치하지 않는 경우가 많습니다. 사실, 살아있는 유기체에는 2 개 이상의 유전자 쌍에 의해 결정되는 많은 특성과 특성이 있으며, 그 반대의 경우도 하나의 유전자가 종종 많은 특성을 제어합니다. 또한, 유전자의 효과는 다른 유전자의 근접성 및 환경 조건에 의해 변경 될 수 있습니다. 따라서, 신생에서, 개별 유전자는 작용하지 않지만, 전체 유전자형은 그 구성 요소 사이의 복잡한 결합 및 상호 작용을 갖는 통합 시스템으로서 작용한다. 이 시스템은 역동적이다 : 돌연변이, 새로운 염색체 및 심지어 새로운 게놈의 결과로 새로운 대립 유전자 또는 유전자의 출현은 시간이 지남에 따라 유전자형의 현저한 변화를 초래한다.
시스템으로서 유전자형의 구성에서 유전자의 작용의 특성은 다양한 상황 및 이하에서 변할 수있다 영향 다양한 요인. 우리가 유전자의 특성과 발현의 특징을 징후로 고려하면 쉽게 볼 수 있습니다.
유전자는 그 작용이 불 연속적입니다. 즉, 다른 유전자와는 그 활성이 분리되어 있습니다.
유전자는 발현에 특이 적입니다. 즉, 유기체의 엄격하게 정의 된 특성이나 성질을 담당합니다.
유전자는 점진적인 방식으로 작용할 수 있는데, 즉 우성 대립 유전자의 수 (유전자 선량)가 증가함에 따라 형질의 발현 정도를 향상시킬 수 있습니다.
단일 유전자는 다양한 특성의 발달에 영향을 줄 수 있습니다. 이것은 유전자의 다중 또는 다발성 작용입니다.
다른 유전자는 동일한 특성 (종종 정량적 특성)의 발달에 동일한 영향을 미칠 수 있습니다. 이들은 여러 유전자 또는 폴리 유전자입니다.
유전자는 다른 유전자와 상호 작용하여 새로운 특성이 나타납니다. 이러한 상호 작용은 제어하에 합성 된 반응의 생성물을 통해 간접적으로 수행됩니다.
유전자의 작용은 염색체상의 위치를 \u200b\u200b바꾸거나 (포지션 효과) 또는 다양한 환경 요인의 작용에 의해 변형 될 수 있습니다.
대립 유전자 유전자 상호 작용. 여러 유전자 (대립 유전자)가 한 특성을 담당하는 현상을 유전자의 상호 작용이라고합니다. 이것이 같은 유전자의 대립 유전자라면, 그러한 상호 작용을 대립 다른 유전자의 대립 유전자의 경우- 비 대립적.
지배적, 불완전한 지배, 과도한 지배 및 코딩 : 대립 유전자 상호 작용의 주요 유형이 구별됩니다.
지배 한 유전자의 두 대립 유전자의 상호 작용 유형 중 하나는 다른 유전자의 행동의 징후를 완전히 배제합니다. 이러한 현상은 다음 조건 하에서 가능하다 : 1) 이종 접합 상태의 우세한 대립 유전자는 모체 형태의 지배적 동형 접합 상태에서와 동일한 품질의 특성을 나타내기에 충분한 생성물의 합성을 제공한다; 2) 열성 대립 유전자가 완전히 비활성이거나 그 활성의 생성물이 우세한 대립 유전자의 활성 생성물과 상호 작용하지 않는다.
대립 유전자 유전자의 이러한 상호 작용의 예는 자주색 지배적이다. 채색 완두콩 꽃 위의 흰색, 부드러운 모양의 씨앗 위의 주름진, 검은 머리 위의 빛, 갈색 눈 위의 인간 등
불완전한 지배 또는 상속의 중간 특성 하이브리드 (이종 접합체)의 표현형이 두 모성 동형 접합체의 표현형과 다를 경우, 즉 형질의 발현이 하나 또는 다른 부모에 대한 편차가 크거나 작은 중간에 관찰된다. 이 현상의 메카니즘은 열성 대립 유전자가 비활성이고, 우성 대립 유전자의 활성 정도가 지배적 특성의 원하는 수준의 발현을 제공하기에 불충분하다는 것이다.
불완전한 지배의 예는 상속입니다. 채색 야행성 식물의 꽃 (그림 3.5). 도표에서 볼 수 있듯이 동형 식물은 빨간색이거나 (AA) 어느 쪽이든 (aa) 꽃과 이형 접합 (Aa) -핑크 F1에서 붉은 꽃이있는 식물과 흰색 꽃이있는 식물을 교차하면 모든 식물에 분홍색 꽃이 생겨 관찰됩니다. 상속의 중간 특성. 하이브리드를 횡단 할 때 와 함께 꽃의 분홍색 채색 F 2 지배적 인 homozygote 때문에 표현형과 유전자형에 의한 절단의 우연의 일치가 있습니다 (AA) 이종 접합체와 다른 (Aa). 따라서 밤의 아름다움을 가진 식물로 간주되는 예에서 F 2 꽃의 색은 일반적으로 다음과 같습니다-1 빨간색 (AA) : 2 분홍색 (Aa) : 1 화이트 (aa).
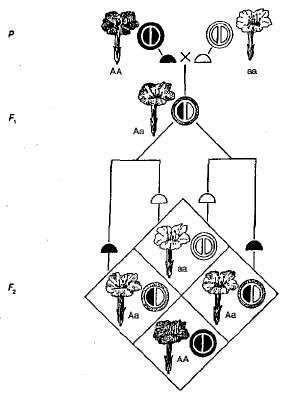
그림. 3.5. 밤의 아름다움에서 불완전한 지배력을 가진 꽃의 색상 상속.
불완전한 지배는 널리 퍼져있는 것으로 입증되었습니다. 인간의 곱슬 머리 상속, 소의 색, 닭의 깃털 색소 및 식물, 동물 및 인간의 다른 많은 형태 적 및 생리적 특성에서 관찰됩니다.
과장 -이형 접합 개체에서 형질의 강한 발현 (Aa) 동형 접합체보다 (AA 그리고 aa). 이 현상은 이종의 근원이라고 가정합니다 (3.7 참조).
코다 미 네이션-이형 접합 개체의 형질 결정에 두 대립 유전자의 참여. 생생하고 잘 연구 된 코딩의 예는 인간의 네 번째 혈액 그룹 (그룹 AB)의 상속 일 수 있습니다.
이 그룹의 사람들의 적혈구에는 두 가지 유형의 항원이 있습니다. A (염색체 중 하나에 존재하는 유전자에 의해 결정) 및 항원 안으로 (또 다른 상동 염색체에 위치한 유전자 / a에 의해 결정). 이 경우에만 두 대립 유전자가 효과를 나타냅니다. 1 A (에서 동형 접합 상태는 혈액 그룹 II, 그룹 A)를 제어하고 나 B (동형 접합 상태에서는 혈액형 III, B 군을 조절합니다). 대립 유전자 1 A 그리고 나 B 서로 독립적으로 이종 접합체에서 작동합니다.
상속 예 그룹 피가 보여줍니다 여러 대립 유전자 : 유전자 /는 세 가지 대립 유전자로 표현할 수 있지만 수십 개의 대립 유전자를 가진 유전자가 있습니다. 한 유전자의 모든 대립 유전자를 여러 대립 유전자의 시리즈, 각 이배체 유기체는 두 개의 대립 유전자를 가질 수 있습니다 (그리고 만). 이들 대립 유전자 사이에서, 나열된 대립 유전자 상호 작용의 모든 변형이 가능하다.
다중 대립 유발 현상은 사실상 일반적입니다. 곰팡이 수정, 종자 식물의 수분, 동물의 머리카락 색 등을 결정하는 동안 호환성의 유형을 결정하는 광범위한 일련의 다중 대립 유전자가 알려져 있습니다.
비 대립 유전자 상호 작용 비 대립 유전자 상호 작용은 많은 식물과 동물에서 설명되어왔다. 그들은 표현형에 따라 비정상적인 쪼개짐의 이종 접합체의 자손으로 나타납니다 : 9 : 3 : 4; 9 : 6 : 1; 13 : 3; 12 : 3 : 1; 15 : 1, 즉 일반적인 멘델 식 공식 9 : 3 : 3 : 1의 수정. 2 개, 3 개 이상의 비 대립 유전자의 상호 작용 사례가 알려져있다. 그 중에서도, 상보성, 전이 및 중합과 같은 주요 유형을 구별 할 수 있습니다.
보완 또는 추가 비 대립적 우성 유전자의 이러한 상호 작용이 호출되며, 그 결과 양쪽 부모에없는 표시가 나타납니다. 예를 들어 두 종류의 스위트 피가 흰 꽃과 교차하면 자주색 꽃을 가진 자손이 나타납니다. 한 종류의 유전자형을 지정하면 AAbb, 그리고 다른- aaBB, 그때
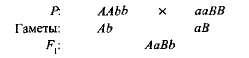
두 개의 주요 유전자를 가진 1 세대 하이브리드 (A 그리고 C) 보라색 안료 안토시아닌 생산에 대한 생화학 적 기초를 얻었지만 단일 유전자는 아님 아 어느 유전자 B도이 안료의 합성을 제공하지 않았다. 안토시아닌 합성은 몇몇 비 대립 유전자에 의해 제어되고, 적어도 2 개의 우성 유전자만으로 제어되는 순차적 생화학 반응의 복잡한 사슬이다 (A-B-) 자주색이 발생합니다. 다른 경우 (aaB- 그리고 A-BB) 식물의 꽃은 흰색입니다 (유전형 공식의“-”표시는 우성 대립 유전자와 열성 대립 유전자가 모두 가능하다는 것을 나타냅니다).
달콤한 완두콩 식물의 자기 수분으로 F 1 안으로 F 2 자주색과 흰 꽃으로 갈라지는 것은 9 : 7에 가까운 비율로 관찰되었습니다. 보라색 꽃이 발견되었다 9/1 6/7, 흰색-7/16 Pennet 격자는이 현상의 원인을 명확하게 보여줍니다 (그림 3.6).
간극 -이것은 한 유전자의 대립 유전자가 다른 유전자의 대립 유전자 쌍의 발현을 억제하는 유전자 상호 작용의 한 유형입니다. 유전자 다른 유전자의 억제 효과를 전이성, 억제제 또는 서프 레서. 억제 된 유전자를 hypostatic.
dihybrid 절단 중 표현형 및 일부 클래스의 수와 비율의 변화에 \u200b\u200b따르면 F 2 여러 유형의 전이 상호 작용을 고려하십시오 : 지배적 전이 (A\u003e B 또는 B\u003e A) 12 : 3 : 1의 절단으로; 열성 전이 (a\u003e B 또는 b \u003e A), 이것은 9 : 3 : 4 분할 등으로 표현됩니다.
폴리머 리아 하나의 표시가 아래에 형성된다는 사실에서 나타남 영향 동일한 표현형 발현을 갖는 몇몇 유전자. 이 유전자들은 폴리머. 이 경우 형질의 발달에 대한 유전자의 독특한 작용 원리가 채택되었습니다. 예를 들어, F 1에서 삼각형 및 타원형 과일 (깍지)이있는 양치기 가방 식물을 가로 지르면 삼각형 모양의 과일이있는 식물이 형성됩니다. 자기 수분으로 F 2 15 : 1의 비율로 삼각형 및 타원형 꼬투리를 갖는 식물로 분할하는 것이 관찰된다. 독특하게 행동하는 두 개의 유전자가 있기 때문입니다. 이 경우 그들은 동일하게 표시됩니다 A 1 그리고 A 2 .
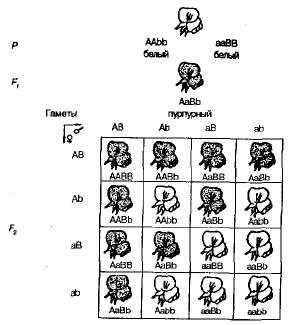
그림. 3.6 . 완두콩의 꽃 채색 상속
그런 다음 모든 유전자형 (A 1 ,-A 2 ,-, A 1 a 2 그러나 2 , 1 a 1 A 2 -) 같은 표현형을 가질 것입니다-삼각 포드와 식물 만 그러나 1 그러나 1 그러나 2 a 2 타원형 포드 모양이 다릅니다. 이 경우입니다 비누 적 중합체.
고분자 유전자는 유형별로 작용할 수 있습니다 누적 중합체. 신체의 유전자형에서 그러한 유전자가 많을수록 유전자 형질의 증가와 함께이 특성의 발현이 강해집니다. (A 1 A 2 A 3 등)의 조치가 요약되거나 누적됩니다. 예를 들어, 밀 곡물 내배의 색 강도는 삼 종교에서 서로 다른 유전자의 우세한 대립 유전자의 수에 비례합니다. 가장 착색 된 곡물 A 1 A 1 A 2 A 2 A 3 , A 3 그러나 곡물 1 그러나 1 그러나 2 a 2 그러나 3 그러나 3 안료가 없었어요
누적 폴리머의 유형에 따라 많은 특성이 상속됩니다 : 우유 생산, 계란 생산, 무게 및 기타 농장 동물의 특성; 사람의 체력, 건강 및 정신 능력의 많은 중요한 매개 변수; 곡물의 스파이크 길이; 사탕무 또는 지질의 뿌리에있는 설탕 함량 해바라기 씨 등
따라서, 다수의 관찰에 따르면, 대부분의 특성의 발현은 상호 작용하는 유전자 복합체 및 환경 조건이 각각의 특정 특성의 형성에 미치는 영향의 결과라는 것을 나타낸다.
유전자 상호 작용
유전자와 형질의 관계는 매우 복잡합니다. 신체에서 하나의 유전자가 항상 하나의 특성만을 정의하는 것은 아니며, 반대로 하나의 특성은 하나의 특성만으로 결정됩니다. 더 자주, 하나의 유전자는 한 번에 여러 특성의 발현에 기여할 수 있으며 그 반대도 마찬가지입니다. 유기체의 유전자형은 독립적 인 유전자의 단순한 합으로 간주 될 수 없으며, 각각은 서로 독립적으로 기능합니다. 하나 또는 다른 특성의 표현형 징후는 많은 유전자의 상호 작용의 결과입니다.
다중 유전자 작용 (다양성)-하나의 유전자가 여러 문자의 형성에 미치는 영향의 과정.
예를 들어, 인간의 경우 머리카락의 붉은 색을 결정하는 유전자는 피부가 더 밝아지고 주근깨가 생깁니다.
때로는 형태 학적 특성을 결정하는 유전자가 생리 기능에 영향을 미쳐 활력과 다산을 줄이거 나 치명적입니다. 따라서 밍크의 푸른 색을 일으키는 유전자는 생식 능력을 감소시킵니다. 동형 접합 상태의 아스트라한 양에서 회색의 우세한 유전자는 상세하다. 그러한 양에서는 위가 저개발되어 잔디 영양으로 전환 할 때 죽기 때문이다.
유전자의 상보적인 상호 작용. 여러 유전자가 한 특성의 발달에 영향을 줄 수 있습니다. 하나의 형질의 발달로 이어지는 몇몇 비 대립 유전자의 상호 작용을 상보 적이라고합니다. 예를 들어, 닭은 네 가지 형태의 빗을 가지고 있으며, 그 중 어느 것의 발현은 두 쌍의 비 대립 유전자의 상호 작용과 관련이 있습니다. 분홍색 유사 문장은 한 대립 유전자의 주요 유전자의 작용에 의해 발생하며 완두 유사 문장은 다른 대립 유전자의 주요 유전자에 기인합니다. 하이브리드에서, 2 개의 우세한 비-대립 유전자가 존재하는 경우, 너트 형 크레스트가 형성되고, 모든 우세한 유전자가없는 경우, 즉 2 개의 비 대립 유전자에 대한 열성 동형 접합체에서, 간단한 융기 부가 형성된다.
유전자의 상호 작용의 결과는 개, 생쥐, 말의 외투 착색, 호박 모양, 완두콩 꽃의 색입니다.
중합은 형질의 발달 정도가 지배적 유전자의 총 수에 의존 할 때 비 대립 유전자의 상호 작용이다. 이 원칙에 따르면 귀리와 밀 곡물의 색과 사람의 피부 색이 상속됩니다. 예를 들어, 비 대립 유전자 두 쌍의 검은 색에는 4 개의 우성 유전자가 있으며, 하얀 피부를 가진 사람은 하나가 아니라 모든 유전자가 열성입니다. 서로 다른 수의 우성 및 열성 유전자의 조합으로 인해 피부색의 다른 강도 (어두운 것에서 밝은 것까지)로 혹 백혈구가 형성됩니다.
유전자 상호 작용의 두 가지 주요 그룹이 있습니다 : 대립 유전자 유전자 사이의 상호 작용과 비 대립 유전자 사이의 상호 작용. 그러나 이것은 유전자 자체의 물리적 상호 작용이 아니라 하나 또는 다른 특성을 결정하는 1 차 및 2 차 제품의 상호 작용이라는 것을 이해해야합니다. 세포질에서 단백질-효소에 의해 유전자에 의해 결정되는 효소 또는 이들 효소의 영향으로 형성된 물질 사이에 상호 작용이 일어난다.
다음과 같은 유형의 상호 작용이 가능합니다.
1) 특정 형질의 형성은 2 개의 효소의 상호 작용을 필요로하며, 그 합성은 2 개의 비 대립 유전자에 의해 결정된다;
2) 하나의 유전자의 참여로 합성 된 효소는 다른 비 대립 유전자에 의해 형성된 효소의 작용을 완전히 억제하거나 비활성화시킨다;
3) 2 개의 효소, 그 형성은 하나의 형질 또는 하나의 과정에 영향을 미치는 2 개의 비 대립 유전자에 의해 제어되어 이들의 조합 된 작용은 형질의 발현의 출현 및 강화로 이어진다.
대립 유전자 유전자 상호 작용
상동 염색체에서 동일한 (상 동성) 유전자좌를 차지하는 유전자를 대립 유전자라고합니다. 각 유기체에는 두 개의 대립 유전자가 있습니다.
대립 유전자 유전자 사이의 이러한 상호 작용 형태는 완전 지배, 불완전한 지배, 코딩 및 과도 배제로 알려져있다.
상호 작용의 주요 형태는 완전한 지배력이며, G. Mendel이 처음 설명했습니다. 그것의 본질은 이형 접합 유기체에서, 대립 유전자 중 하나의 발현이 다른 대립 유전자의 발현을 지배한다는 사실에 있습니다. 1 : 2 : 1 유전자형 절단의 완전한 지배로, 그것은 3 : 1의 표현형 절단과 일치하지 않습니다. 의학 실습에서 2 천 개의 단발성 유전병에서 거의 절반이 정상 유전자보다 병리학 적 유전자가 지배적으로 나타납니다. 이종 접합체에서, 병리학 적 대립 유전자는 대부분 질병의 징후 (주요 표현형)에 의해 나타난다.
불완전한 지배는 이형 접합 유기체 (Aa)에서 지배적 인 유전자 (A)가 열성 유전자 (a)를 완전히 억제하지 않아 상호 작용의 형태로 부모의 특성 사이에 중간체가 나타납니다. 여기서 유전자형과 표현형에 의한 분리는 일치하고 1 : 2 : 1에 이릅니다.
이형 접합성 유기체에서 코딩 할 때, 각각의 대립 유전자 유전자는 그것에 의존하는 생성물, 즉 두 대립 유전자의 생성물이 형성되게한다. 이 증상의 전형적인 예는 인간 적혈구 세포가 표면에 항원을 가지고있는 두 대립 유전자에 의해 혈액 그룹 시스템, 특히 ABO 시스템이다. 이러한 형태의 표현을 코딩이라고합니다.
과잉 지배-이형 접합 상태의 우성 유전자가 동형 접합 유전자보다 더 두드러 질 때. 따라서 Drosophila에서는 AA 유전자형을 사용하여 정상적인 수명을 기대할 수 있습니다. Aa — 삶의 연장 된 삼자주의; AA-죽음.
다원주의
각 유기체에는 단 2 개의 대립 유전자가 있습니다. 그러나, 일부 유전자좌가 다른 상태에있을 수있는 경우, 사실상 대립 유전자의 수는 종종 2 개 이상일 수있다. 그러한 경우에, 그들은 다수의 대립 유전자 또는 다수의 대립 유전자에 대해 말한다.
다수의 대립 유전자는 상이한 지수를 갖는 동일한 문자로 표시된다 : 예를 들어 : A, A1, A3 ... 대립 유전자 유전자는 동종 염색체의 동일한 영역에 위치한다. 2 개의 상동 염색체가 핵형에 항상 존재하기 때문에, 다수의 대립 유전자를 사용하여 각 유기체는 한 번에 2 개의 동일하거나 상이한 대립 유전자를 가질 수있다. 그들 중 하나만이 생식 세포에 들어갑니다 (동종 염색체의 차이와 함께). 다수의 대립 유전자에 대해, 동일한 형질에 대한 모든 대립 유전자의 특징적인 효과. 그들 사이의 차이점은 부호의 발달 정도에만 있습니다.
두 번째 특징은 체세포 또는 이배체 유기체의 세포가 염색체의 동일한 유전자좌에 위치하기 때문에 최대 두 개의 대립 유전자를 포함한다는 것입니다.
다수의 대립 유전자에 내재 된 또 다른 특징. 지배의 본질에 의해, 대립 형질 문자는 순차적 인 순서로 배열된다 :보다 일반적이고 변하지 않는 문자가 다른 문자를 지배하고, 시리즈의 두 번째 유전자는 첫 번째에 비해 열성이지만, 다음을 지배한다. 인간에서 다수의 대립 유전자의 발현의 한 예는 ABO 시스템의 혈액 그룹이다.
다중 에일 리즘은 조합 가변성, 특히 유전자형을 향상시키기 때문에 생물학적으로 실질적으로 중요하다.
비단 일 유전자 상호 작용
특성이나 특성이 서로 상호 작용하는 둘 이상의 분리 불가능한 유전자에 의해 결정되는 경우가 많이 있습니다. 비록 유전자가 상호 작용하는 것이 아니라 그들이 제어하는 \u200b\u200b제품이기 때문에 상호 작용은 조건부입니다. 이 경우 Mendelivian 분할 법칙에서 벗어납니다.
유전자 상호 작용의 4 가지 주요 유형이 있습니다 : 상보성, 전이, 중합 및 변형 작용 (다양성).
상보성은 하나의 우성 유전자가 다른 비-유전자 우성 유전자의 작용을 보완하고 함께 부모에없는 새로운 특성을 정의 할 때 비-대립 유전자의 상호 작용의 한 유형입니다. 또한, 상응하는 형질은 비 대립 유전자 둘 다의 존재 하에서 만 발생한다. 예를 들어, 생쥐에서 코트의 유황 색은 두 유전자 (A와 B)에 의해 제어됩니다. 유전자 A는 안료의 합성을 결정하지만, 동형 접합체 (AA) 및 이형 접합체 (Aa)는 모두 알비노이다. 다른 유전자 B는 주로 모발의 기저 및 말단에 안료 축적을 제공한다. digeterozygotes (AabB x AabB)의 교차는 9 : 3 : 4의 비율로 하이브리드의 절단을 초래합니다. 상보 적 상호 작용에 대한 수치 비율은 최대 9 : 7 일 수 있습니다. 9 : 6 : 1 (Menlivliv 분할의 수정).
인간에서 상보적인 유전자 상호 작용의 예는 보호 단백질 인 인터페론의 합성입니다. 신체의 형성은 다른 염색체에 위치한 두 개의 비 대립 유전자의 상보 적 상호 작용과 관련이 있습니다.
간질은 하나의 유전자가 다른 비 대립 유전자의 작용을 억제하는 비 대립 유전자의 상호 작용입니다. 우성 및 열성 유전자 (A\u003e B, a\u003e B, B\u003e A, B\u003e A)는 모두 억제를 유발할 수 있으며, 이에 따라 전이가 우성 및 열성으로 식별됩니다. 억제 유전자를 억제제 또는 억제제라고합니다. 억제제 유전자는 일반적으로 특정 형질의 발달을 결정하지는 않지만 다른 유전자의 작용 만 억제합니다.
효과가 억제 된 유전자를 hypostatic이라고합니다. 전이성 유전자 상호 작용으로, F2에서의 표현형 절단은 13 : 3이고; 12 : 3 : 1 또는 9 : 3 : 4 기타 호박 과일의 색상과 말의 색상은 이러한 유형의 상호 작용에 의해 결정됩니다.
 가요? 차이점은 무엇입니까
가요? 차이점은 무엇입니까 우리는 Ba Zi 카드에서 출생 시간을 결정합니다- "삶은 좋다"-풍수에 따르면 당신의 삶에 대한 긍정적 인 생각
우리는 Ba Zi 카드에서 출생 시간을 결정합니다- "삶은 좋다"-풍수에 따르면 당신의 삶에 대한 긍정적 인 생각 시리우스는 지구에 큰 영향을 미쳤으며 계속 영향을 미치고 있습니다.
시리우스는 지구에 큰 영향을 미쳤으며 계속 영향을 미치고 있습니다.