Az emberi genetika fejlődési szakaszainak története rövid. A genetika történetének főbb időpontjai
bevezetés
A biológiai rendszerek képesek olyan információkat tárolni és továbbítani olyan struktúrák és funkciók formájában, amelyek a múltban a hosszú evolúció eredményeként keletkeztek.
Felfedeztek olyan mobil genetikai elemeket, amelyek olyan általános biológiai jelenségekben érintettek, mint a nitrogén rögzítése, rosszindulatú sejtnövekedés, az immunrendszer munkája és a baktériumok antibiotikumokhoz való alkalmazkodása, instabil mutációk és anyai öröklődés.
A genetika a szervezetek öröklődésének és variabilitásának és kezelési módszereinek biológiai tudománya.
A genetika központi fogalma a "gén". Ez az öröklődés elemi egysége, amelyet számos jel jellemez.
A genetika alapját az öröklési törvények állapították meg, amelyeket G. Mendel osztrák biológus fedezett fel, amikor számos kísérletet végzett különböző borsófajták átkelésében.
A kutatók főbb irányai - genetikusok:
1. Olyan nukleinsavmolekulák tanulmányozása, amelyek az egyes élő fajok genetikai információi, az öröklési egységek felügyelői.
2. A genetikai információ továbbításának mechanizmusainak és mintáinak vizsgálata.
3. A genetikai információ megvalósításának mechanizmusainak tanulmányozása az élő állatok sajátos jeleiben és tulajdonságaiban.
4. A genetikai információ változásának okainak és mechanizmusainak megvilágítása a szervezet fejlődésének különböző szakaszaiban.
\u003e A genetika mint tudomány fejlődésének története
Charles Darwin tanításának széles körű elterjedése után az egyik első kritikus, aki elméletileg gyenge pontot mutatott, a skót kutató, F. Jenkins. 1867-ben megjegyezte, hogy a darwini elméletben nincs egyértelműség, hogy miként halmozódik fel az utódok változásai. Elvégre először csak bizonyos egyénekben fordulnak elő változások. A normál egyénekkel való átkelés után nem kell megfigyelni a felhalmozódást, hanem ezt a tulajdonságot az utódokban hígítani. Azaz az első generáció marad? a második változás? változások, stb. a funkció teljes eltűnéséig. Charles Darwin nem talált választ erre a kérdésre.
Eközben megoldást találtak erre a kérdésre. Brno (Cseh Köztársaság) G. Mendel szerzetesi iskolájának tanárát kapta. 1865-ben közzétették a borsófajták hibridizációjával kapcsolatos munkájának eredményeit, ahol felfedezték az öröklés legfontosabb törvényeit. A szerző kimutatta, hogy a szervezetek jellemzőit diszkrét örökletes tényezők határozzák meg.
Még Charles Darwin könyve közzététele előtt még a hibridek különböző nemzedékeiben a genotípusok változásának sorsát kívánta nyomon követni. A vizsgálat célja a borsó volt. Mendel kétféle borsót vitt - sárga és zöld magokkal. Miután átlépte a két fajtát, a hibridek első generációjában csak sárga magokkal rendelkező borsót talált. Az általa kapott hibridek beporzásával megkapta a második generációt. A zöld magokkal rendelkező személyek megjelentek benne, de szignifikánsan kisebbek voltak, mint a sárga. A Mendel arra a következtetésre jutott, hogy ezek száma és mások száma megszámlálásra került, hogy a sárga magokkal rendelkező egyének száma a zöldben élő egyének számát jelenti, kb. 3: 1.
Ezzel párhuzamosan több kísérletet is végzett növényekkel, több generációban nyomon követve a jeleket. Az első generáció minden tapasztalatában csak a szülői jelek egyike jelent meg. Mendel dominánsnak nevezte. Ideiglenesen eltűnő tulajdonsága, recesszívnek hívta. Minden kísérletben a domináns jellemzőkkel rendelkező egyedek száma a második generációs hibridek körében recesszív tulajdonságú egyedek számával átlagosan 3: 1 volt.
Tehát azzal érvelhetünk, hogy az ellentétes tulajdonságokkal rendelkező növények átlépésekor nem a tulajdonságok hígítása, hanem az egyik tulajdonság elnyomása egy másik által, ezért meg kell különböztetni a domináns és a recesszív tulajdonságokat.
Mendel tovább ment a kísérleteiben. A második generáció önszennyezett hibridjeit és a harmadik és a negyedik generáció hibridjeit kapta. Megállapította, hogy a második generáció recesszív tulajdonságú hibridjei a további reprodukció során nem oszlanak meg a harmadik vagy a negyedik generációban. A domináns második generációs hibridek egyharmada ugyanúgy viselkedik. A domináns hibridek kétharmada a harmadik generációs hibridre váltáskor szétválik, ismét 3: 1 arányban. A harmadik generációs hibridek, amelyek recesszív tulajdonságokkal rendelkeznek ebből az elválasztásból, és a hibridek egyharmada a negyedik generációra való áttérés során nem oszlanak meg, és a fennmaradó harmadik generációs hibridek megoszlanak, és ismét 3: 1 arányban.
Ez a tény fontos tényt mutat: az azonos külső jelekkel rendelkező egyének különböző örökletes tulajdonságokkal rendelkezhetnek, azaz a fenotípus szerint nem lehet eléggé teljességgel megítélni a genotípust. Ha az egyén nem észleli az osztódást az utódokban, akkor azt homozigótanak nevezik, ha igen, heterozigóta.
Ennek eredményeként G. Mendel megfogalmazta az első generáció hibridjeinek egységességének törvényét: a hibridek első generációja, amely csak a domináns jellemzők megjelenése miatt van, mindig egyenletes. Ezt a törvényt Mendel első törvényének vagy az uralom törvényének is nevezik. Kutatásának eredményei azonban szinte 35 évig - 1865 és 1900 között - gyakorlatilag ismeretlenek voltak.
1900-ban Mendel törvényeit három tudós önállóan újra felfedezte, G. de Vries Hollandiában, C. Correns Németországban és E. Cermak Ausztriában. 1909-ben a dán tudós, V. Iogansen bevezette a "gén" fogalmát (a görög "eredet" szóból).
Az 1910-1915-ben A. Weisman, T. Morgan, A. Stertevant, G. J. Meller és mások írásaiban kifejlesztett öröklési kromoszómaelmélet azt állítja, hogy a test tulajdonságainak és tulajdonságainak átruházása generációról generációra (öröklődés) elsősorban a kromoszómák, amelyekben gének találhatók.
1944-ben az amerikai biokémikusok (O. Avery és mások) megállapították, hogy a DNS az öröklési tulajdonság hordozója.
Ettől kezdve elkezdődött a tudomány gyors fejlődése, feltárva az élet fő megnyilvánulásait molekuláris szinten. Ugyanakkor először megjelent a tudomány új fogalma - a molekuláris biológia. A molekuláris biológia azt vizsgálja, hogyan és milyen mértékben határozza meg a biológiailag fontos molekulák (főként fehérjék és nukleinsavak) szerkezetét és tulajdonságait az élőlények növekedése és fejlődése, az örökletes információk tárolása és továbbítása, az élő sejtekben az energia átalakulása és más jelenségek.
1953-ban a DNS-struktúrát megfejtettük (F. Crick, D. Watson). A DNS szerkezetének megfejtése azt mutatta, hogy a DNS molekula két komplementer polinukleotid láncból áll, amelyek mindegyike mátrixként szolgál új analóg láncok szintéziséhez. A DNS megkétszerező tulajdonsága biztosítja az öröklődés jelenségét.
A DNS szerkezetének megfejtése a molekuláris biológia forradalma volt, amely fontos felfedezések időszakát nyitotta meg, amelynek általános iránya az élet lényegéről, az öröklődés természetéről, a variabilitásról és az anyagcseréről szóló ötletek kidolgozása volt. A molekuláris biológia szerint a fehérjék nagyon összetett makromolekulák, amelyek szerkezeti elemei aminosavak. A fehérje szerkezetét az aminosavak szekvenciája határozza meg. Ugyanakkor, az aminosavak szerves kémiájából ismert 100-ból mindössze húszat használnak az összes szervezet fehérjéinek kialakulásában. Még mindig nem világos, hogy ezek a 20 aminosav szintetizálják a szerves világ fehérjéit. Általában minden olyan teremtményben, amely a Földön él, 20 aminosav, 5 bázis, 2 szénhidrát és 1 foszfát található.
A XIX. Század végére a mikroszkópok optikai tulajdonságainak növelése és a citológiai módszerek javítása révén lehetővé vált a kromoszómák viselkedésének megfigyelése a gametákban és a zigótákban.
Az öröklődés anyagi alapjai mintegy 50 évvel ezelőtt kezdtek tisztázni, amikor F. Crick és J. Watson megfejtette a DNS szerkezetét. Ezt megelőzően a biológusok, akik az örökletes tulajdonságok átadását tanulmányozták, átléptek, rájöttek, hogy minden tulajdonságot egy külön részecske határoz meg, amelyet génnek neveztek. Kiderült, hogy a gének a sejtmagban vannak, a kromoszómákban. A DNS szerepének és a fehérjeszintézis mechanizmusának felfedezése után világossá vált, hogy a gén a DNS-lánc egy része, amelyen egy bizonyos fehérje molekula szerkezetét rögzítik. Néhány génben csak 800 nukleotid pár, másokban mintegy egymillió. Emberekben csak mintegy 90 ezer gén.
A DNS-molekulák mindegyik szakasza négyféle egységből álló lánc - egy nukleotid, egy másik sorrendben. A nukleotidokat rendszerint pároknak tekintjük, mivel a DNS molekulában két lánc és nukleotidja párhuzamosan összekapcsolódik. Négy nukleotidfajta, négy "betű" lehetővé teszi, hogy írjon egy genetikai szöveget, amelyet az élő sejtekben a fehérjeszintézis mechanizmusai olvasnak. A három egymást követő nukleotid egy csoportja, amely egy viszonylag komplex transzmissziós mechanizmuson keresztül hat, a fehérje-szintézisben résztvevő intracelluláris részecskéket okozó riboszómát egy bizonyos aminosav felvételére a citoplazmából okozza. Ezután a következő három nukleotid a közvetítőkön keresztül diktálja a riboszómának, hogy melyik aminosav a fehérje láncba kerül a következő helyre, és így a fehérje molekula fokozatosan nyerhető. A DNS-ben a nukleotidpárok háromszorosával rögzített információ elegendő ahhoz, hogy egy új szervezetet építsünk be minden jellemzőjével.
A genetikai információt nukleotidszekvencia formájában tároljuk. A DNS-ből az RNS-be továbbítja a sejtet. Ennek a reakciónak a során a DNS-szekvencia egy részét reprodukáljuk, a gént szintetizáljuk és a messenger-RNS-t szintetizáljuk. A csak egy szálból álló hírvivő RNS szekvenciája komplementer az azt kódoló DNS-szál nukleotidszekvenciájához.
Új genetikai ág született - genomika, amely a teljes genomokat tanulmányozza. Egészen a közelmúltig, a molekuláris biológia és a géntechnológia előrehaladása alapján, meg lehetett olvasni a gombák, élesztőbaktériumok vírusainak genetikai szövegét, és végül 1998-ban, 8 év kemény munka után el tudtuk olvasni a többsejtű állat, egy nematódus (egy talajban élő kis féreg) genomját. Megfejtette az emberi genomot. A nematód genom körülbelül 100 millió nukleotidpárból áll. Az emberi genom 3 milliárd párból áll. Létrehozta a "Human Genome" nemzetközi programot. A különböző országokban működő laboratóriumok a nukleotid (szekvenálás) adatok dekódolását jelentik a nemzetközi adatbázisban, amely minden kutató számára elérhető.
Eredményei elengedhetetlenek az emberek és más fajok eredetének megértéséhez, a molekulák és sejtek fejlődéséhez, az információ kölcsönhatásához az anyagok áramlásához és az élő rendszerek energiájához. Ma a tudósok teljesen megfejtették az emberi testben jelen lévő összes gén szerkezetét és helyét. De a gének működésének törvényei megértéséhez jelentős időre és pénzre van szükség - egy olyan pontszám, amely a szólistákat (géneket) harmonikus zenekarokká alakítja.
Az E. Haeckel 1866-ban fogalmazta meg a Biogenetikai törvényt C. Darwin elképzelései és F. Muller kutatása alapján. Ezért az úgynevezett E. Haeckel-F. Muller biogenetikai törvény. A biogenetikai törvényt 1910-ben alapvetően N. N. Severtsev (1866 - 1936) tisztázta, aki létrehozta a filembriogenezis elméletét.
E törvény szerint a fejlődési folyamatban lévő embriók valamilyen rövidített formában ismétlődnek az őseik által áthaladó evolúciós úton, vagyis az embrió fejlődése és az evolúciós folyamat között van hasonlóság. Megállapítást nyert, hogy az állatok magasabb formáinak embriói hasonlóak az alsó formák embrióihoz. Az embrió korai fejlődési szakaszai minden gerincesben rendkívül hasonlóak, és nem könnyű megkülönböztetni az emberi embriót a sertés, csirke, béka vagy hal embriójától. A filogenetikai tulajdonságok ontogenezisében az ismétlés (recapitáció) nem teljes, bizonyos további evolúciós transzformációkkal kapcsolatos torzulások, különösen az ősi formák megfelelő fejlődési fázisainak jellemzői megismételhetők.
A biogenezis legfontosabb eredménye a genetikai programozási eszközök kialakítása, amely lehetővé teszi az elért konszolidációt. A különböző programok küzdelme a létező küzdelemben két fontos következményhez vezet:
Először is, a természetes kiválasztás javítja az egyének egyedi fejlesztési programjait.
Másodszor, a fajok fejlődési irányának programozása történik. Ebben az esetben maga a bioszféra lesz a programozó eszköz. Végül is meghatározza az összetételét alkotó fajok evolúciós átalakulásának jellemzőit, sebességét és irányát.
A genetika mint tudomány története nem sokkal régen kezdődött, kb. A 19. század második felétől Gregor Mendel cseh szerzetes munkájával. Ezt megelőzően az emberek nem tudták, mi az alapja az organizmusok öröklődésének és variabilitásának. Ezen a területen nem végeztek szisztematikus kutatást. Ősi idők óta két hipotézis van arról, hogy az utódok miért hasonlítanak az őseikre. Ezek a közvetlen és közvetett öröklés hipotézisei.
A közvetlen öröklés hipotézise szerint azt feltételezték, hogy a szülő testének minden egyes része, mindegyik sejtje átadja szerkezetének sajátosságait a leszármazónak. A közvetett öröklés hipotézise abból a tényből ered, hogy a test nem minden része részt vesz a nemi szervek kialakulásában, és elkülönülve alakulnak ki, így a szülő élettartambeli változásai nem kerülnek utódokra.
Charles Darwin ragaszkodott az első hipotézishez, ami tévedés volt. Emiatt nem tudta elvetni evolúciós elméletének egyik kritikáját, aki az alábbiakat írta. Ha egyetlen állatnak van jele, akkor a következő generációhoz való továbbítással ½ balra, másik ¼-re, stb. Lesz. Végül a jel eltűnik. Darwin nem tudott vitatkozni, mert nem tudta, hogy a test jelei diszkrétek, nem keverednek és nem oldódnak.
G. Mendel hosszú, szisztematikus, statisztikai vizsgálatokat végzett a karakterek örökléséről. A tanulmány tárgya a borsó. Nagyon jó választás volt. Az olyan jellemzők, mint a magok és a virágok színe, a vetőmag-ráncok és a több más kromoszómák (azaz nem öröklődnek), és leggyakrabban nincsenek közbenső megnyilvánulása a tulajdonságnak (például a vetőmag lehet sárga vagy zöld, de nem kevert szín). Mendel természetesen nem tudott semmit a láncolt öröklésről, a gének és allélek egymásra gyakorolt \u200b\u200bhatásáról, stb. Ha mindezt megfigyelték a borsó tanulmányozott jeleiben, akkor kísérlete nem sikerült.
Gregor Mendel és az örökség törvényei megjelentek számukra - 1860-as években
Mendel bizonyította az örökletes vonások diszkrécióját. Nem hígítják őket, és csak egymás elnyomása van. A Mendel hibridológiai kutatási módszert dolgozott ki. De először a genetika történetében Mendel megfogalmazta a legfontosabb dologot: az első generáció hibridjeinek egységességét, a második generációban történő szétválasztást, a független örökséget.
Azonban ezekben a napokban a genetika története még nem kezdődött el. Mendel önképzett szerzetes volt, és kutatása nem nagy jelentőséget tulajdonított. Csak a 20. század elején, amikor sok tudós kísérletesen kísérleteztek a különböző növényekre és állatokra, megerősítette Mendel törvényeinek érvényességét, művei jól megérdemelt értékelést kaptak.
A 20. század eleje a genetika fejlődésének turbulens fázisa volt. Ekkor megjelenik a "genetika" kifejezés. A „gén”, a „genotípus” és a „fenotípus” meghatározását adjuk meg. A gének (társ-öröklés) jelenségét felfedezik, W. Batson megnyitja, stb. 1910-ben T. Morgan más tudósokkal együtt kromoszóma elméletet dolgoz ki, amely nagyrészt összefoglalja és elmagyarázza a genetika történetében előzőleg felfedezett összes felfedezést.
 Thomas Morgan a kromoszómákat tanulmányozta
Thomas Morgan a kromoszómákat tanulmányozta
A következő években a genetika és az evolúciós doktrínák egymáshoz kapcsolódtak. A második az első törvényei szerint magyarázható.
A tudósok tudták, hogy a kromoszómák részt vesznek a genetikai információk továbbításában, de nem tudták, hogy melyik anyag felelős. A 40-es években világossá vált, hogy a DNS öröklési hordozó. Tehát számos tudós átadta néhány baktérium DNS-jét másoknak, és a másodikban megfigyelte az első jelek megjelenését.
A kémia és a fizika fejlődésével lehetővé vált a DNS szerkezetének vizsgálata, amelyet Crick és Watson 1953-ban végzett. Kiderült, hogy a molekula két polinukleotid láncból áll, amelyek spirálba vannak csavarva. A DNS-szálak mindegyike egy új szál-szintézishez használható, amely kiegészíti a DNS-t, és a DNS megduplázása biztosítja az öröklődést.
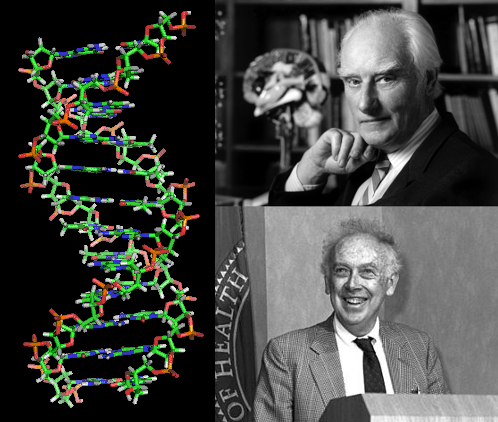 Francis Crick és James Watson felfedezték a DNS szerkezetét - az 1950-es éveket
Francis Crick és James Watson felfedezték a DNS szerkezetét - az 1950-es éveket
A tudósok ezt megértették a gén nukleotidszekvenciája meghatározza a fehérje molekula szerkezetét..
A genetika történetében a 20. század 70-es éveit a géntechnológia fejlődése jellemezte. A tudósok elkezdték zavarni az élő szervezetek genomjait és megváltoztatták őket. Megkezdte a különböző fiziológiai folyamatok molekuláris alapjának tanulmányozását.
A 20. század utolsó évtizedében számos egyszerű organizmus genomját szekvenálták (megfejtették). A XXI. Század elején (2003) befejeződött az emberi genom dekódolásának (a nukleotidok szekvenciájának meghatározása) projektje.
 A "Human Genome" projekt logója
A "Human Genome" projekt logója
Napjainkig számos szervezet adatbázisai léteznek. Egy ilyen személy adatbázisának jelenléte nagy jelentőséggel bír számos betegség megelőzésében és tanulmányozásában.
A genetika születését a két évszázad fordulóján (1900) a biológiai tudomány teljes előzményeinek előkészítése végzi. XIX. két nagy felfedezés miatt lépett be a biológia történetébe: M. Schleiden és T. Schwann (1838) által megfogalmazott sejtelmélet és C. Darwin evolúciós tanításai (1859). Mindkét felfedezés döntő szerepet játszott a genetika fejlődésében. A sejtelmélet, amely a sejtet az összes élőlény fő szerkezeti és funkcionális egységének nyilvánította, fokozott érdeklődést váltott ki annak struktúrájának tanulmányozása iránt, ami tovább vezetett a kromoszómák felfedezéséhez és a sejtosztódási folyamat leírásához. C. Darwin elmélete viszont az élő szervezetek legfontosabb tulajdonságait érinti, amelyek később a genetika tanulmányozásának tárgyát képezték - öröklődés és variabilitás. Mindkét elmélet a XIX. egyesítette azt az elképzelést, hogy szükség van ezeknek a tulajdonságoknak a hordozóira, amelyeknek a cellákban kell lenniük.
A huszadik század elejéig. Az öröklési mechanizmusokra vonatkozó minden hipotézis tisztán spekulatív volt. Így Charles Darwin (1868) pangenesisének elmélete szerint apró részecskék, gemulák, amelyek a véráramban keringenek, és bejutnak a csírasejtekbe, elkülönülnek a test minden sejtjétől. A csírasejtek egyesülése után egy új szervezet kialakulása során minden gemmulumból egy azonos típusú sejtet kaptunk, amelyből eredetileg minden tulajdonság, beleértve a megszerzett szülőket is. Darwin álláspontja a jelek átadásának mechanizmusáról a szülőktől az utódoknak a véren keresztül még mindig az ókori görög filozófusok természetes filozófiájában rejlik, ideértve a Hippokratész tanításait is (Kr. E. 5. század).
Egy másik spekulatív hipotézist az öröklésre 1884-ben terjesztett elő K. Negeli (német). Azt javasolta, hogy az öröklési, idioplazmás, a sejtekben összegyűjtött molekulák nagy filamentus szerkezetekbe, a micellákba tartozó különleges anyag vegyen részt az öröklődő hajlamok utódokba történő átvitelében. A micellák kötegekben vannak összekapcsolva és egy olyan hálózatot alkotnak, amely áthatol minden sejtre. Idioplazmának mind a szexuális, mind a szomatikus sejtjei vannak. A citoplazma többi része nem vesz részt az örökletes tulajdonságok átvitelében. Anélkül, hogy tényeket támasztana alá, K. Negeli hipotézise mindazonáltal elvárta az öröklés anyagi hordozóinak létezéséről és strukturáltságáról szóló adatokat.
Első alkalommal A. Weisman rámutatott a kromoszómákra, mint az öröklődés anyagi hordozóira. Elméletében a német citológus, Wilhelm Roux (1883) következtetéseit az örökletes tényezők (kromatinszemcsék) lineáris elrendezéséről a kromoszómákban és az örökletes anyag eloszlásának lehetséges módjaként az osztódás során fennálló hosszanti kromoszómaelosztást illeti. A. Weisman „csíráinak” elméletét 1892-ben véglegesítették. Úgy vélte, hogy a szervezetben az öröklődés különleges tartalma van - a „csírázás”. A csírasejtek szubsztrátja a csírasejtek kromatinszerkezete. A csírasejtek halhatatlanok, a csírasejteken keresztül, az utódoknak továbbítják, míg a szervezet teste, a soma halandó. A Germplaszm diszkrét részecskékből áll - egy biofórból, amelyek mindegyike a sejtek külön tulajdonságát határozza meg. A bioforokat a sejtek specializációját meghatározó részecskékre csoportosítják. Ezek viszont magasabb rendű (ides) struktúrákba egyesülnek, amelyekből kromoszómák képződnek (A. Weisman terminológiájában).
A. Weisman tagadta a megszerzett tulajdonságok öröklésének lehetőségét. Az örökletes változások forrása az ő tanításai szerint a műtrágyázási folyamat során előforduló események: a csírasejtek érése során bizonyos információk (csökkentés) elvesztése és az apa és az anya meghatározó tényezőinek keverése, ami új tulajdonságok kialakulásához vezet. A. Weisman elmélete hatalmas befolyást gyakorolt \u200b\u200ba genetika fejlődésére, meghatározva a genetikai kutatás további irányát.
A huszadik század elejére. valódi előfeltételeket teremtettek a genetikai tudomány fejlesztésére. A döntő szerepet az 1900-ban újra felfedezett G. Mendel törvényei játszották. Egy cseh amatőr kutató, a Brno-kolostor szerzetese, Gregor Mendel, 1865-ben megfogalmazta az öröklés alapvető törvényeit. Ezt lehetővé tette az első tudományos genetikai módszer kifejlesztése, amelyet „hibridológiai” -nak neveztek. Keresztrendszeren alapult, amely lehetővé teszi a karakterek öröklési mintáinak feltárását. Mendel három törvényt és a „gameták tisztaságát” szabályt fogalmazott meg, amelyet részletesen tárgyalunk a következő előadásban. Nem kevésbé (és talán még inkább) fontos volt, hogy Mendel bevezesse az örökletes hajlamok (gének prototípusai) fogalmát, amely a tulajdonságok fejlődésének anyagi alapja, és ragyogó feltételezést fogalmazott meg a „tiszta” gaméták fúziója következtében történő párosításukról.
Mendel kutatása és nézetei az öröklési mechanizmusról a tudomány fejlődését megelőző évtizedek előtt. Még a fent említett öröklődés természetére vonatkozó spekulatív hipotéziseket később is megfogalmazták. A kromoszómákat még nem fedezték fel, és a sejtosztódás folyamatát, amely az örökletes információk a szülőktől a leszármazottakhoz való továbbításának alapja, nem ismertették. Ebben az összefüggésben a kortársok, még azok is, akik Charles Darwinhoz hasonlóan ismerik G. Mendel munkáit, nem értékelik a felfedezését. A biológiai tudomány 35 évig nem állította azt.
Az igazságosság 1900-ban győzedelmeskedett, amikor Mendel törvényeinek másodlagos újbóli felfedezése után három tudós egyidejűleg és önállóan követte: G. de Vries (cél), C. Correns (őt) és E. Cermak (szülő). Megismételve Mendel kísérleteit, megerősítették az általa felfedezett törvények egyetemes természetét. Mendel kezdett tekinteni a genetika alapítójának, és 1900-ban elkezdte a tudomány fejlődésének visszaszámlálását.
A genetika történetében általában két periódust különböztetünk meg: az első a klasszikus, formális, genetikai időszak (1900-1944) és a második a molekuláris genetika időszaka, amely a mai napig folytatódik. Az első időszak legfőbb jellemzője, hogy az öröklődés anyagi hordozóinak jellege ismeretlen maradt. A dán genetikus V. Johansen, a mendeli örökletes tényező analógja által bevezetett „gén” fogalma absztrakt volt. Itt egy idézet az 1909-es munkájából: „A szervezet tulajdonságait speciális, különálló körülmények között különválasztva, és ezért bizonyos mértékig független csírasejtek, vagyis a géneknek nevezett egységek vagy elemek határozzák meg. Jelenleg a gének természetét nem lehet egyértelműen megérteni, csak akkor lehet elégedettek, ha ilyen elemek léteznek. De kémiai képződmények? Erről még nem tudunk semmit. Annak ellenére, hogy a gén fiziko-kémiai természetéről hiányzik a tudás, ebben az időszakban feltárták a genetika alapvető törvényeit, és kifejlesztették a tudomány alapját képező genetikai elméleteket.
A Mendel törvényeinek újbóli felfedezése 1900-ban vezetett a tanításai gyors terjedéséhez és a különböző országokban a különböző helyeken (csirkék, lepkék, rágcsálók stb.) Végzett kutatók számos kísérletének megerősítéséhez, törvényei egyetemes jellegének megerősítéséhez. Ezen kísérletek során új öröklési mintákat fedeztek fel. 1906-ban W. Batson és R. Pennet angol tudósok a Mendel törvényeiről való első eltérést ismertették, amelyet később géncsatolásnak neveztek. Ugyanebben az évben L. Doncaster, az angol genetikus, egy pillangó kísérletben felfedezte, hogy a jellemzők a padlóhoz kapcsolódnak. Ugyanakkor a huszadik század elején. Megkezdődik a tartós örökletes mutációs változások vizsgálata (G. de Vries, S. Korzhinsky), és megjelennek az első művek a populációk genetikájáról. 1908-ban G. Hardy és V. Weinberg megfogalmazta a populációs genetika alapjogát a génfrekvenciák állandóságán.
A klasszikus genetika időszakának legfontosabb tanulmányai azonban a kiváló amerikai genetikus T. Morgan és tanítványai művei voltak. T. Morgan a világ legnagyobb genetikai iskolájának alapítója és vezetője, ahonnan a tehetséges genetikusok teljes galaxisa jött. Tanulmányaiban Morgan először használta a Drosophila gyümölcslevelet, amely kedvenc genetikai tárgyává vált, és továbbra is most marad. A W. Betson és R. Pennet által felfedezett génkötés jelenségének vizsgálata lehetővé tette a Morgan számára, hogy megfogalmazza az öröklési kromoszóma elmélet főbb pontjait, amelyeket az alábbiakban részletesen tárgyalunk. Ennek az alapvető genetikai elméletnek a fő tézise az volt, hogy a gének lineárisan vannak elrendezve a kromoszómában, mint a szál gyöngyei. Azonban még 1937-ben Morgan azt is elmondta, hogy a genetikusok között nincs egyetértés a gén természetében - függetlenül attól, hogy valóságosak vagy absztrakciók. Megjegyezte azonban, hogy minden esetben a gén egy adott kromoszómához kapcsolódik, és tiszta genetikai analízissel lokalizálható.
Számos más kiemelkedő tanulmányt végeztek Morgan és munkatársai (T. Painter, Bridges, A. Stertevant és mások): a genetikai feltérképezés elve, a nemi meghatározás kromoszóma-elmélete, a polytén kromoszómák szerkezetét vizsgálták.
A klasszikus genetika időszakának fontos eseménye a mesterséges mutagenezisre épülő művek kifejlesztése volt, amelyek első adatait 1925-ben szerezték be a Szovjetunióban, G.A. Nadson és TS Filippov a rádiumos élesztősejtek besugárzásának kísérleteiben. Ebben az irányban a munka fejlesztésének döntő fontosságúak voltak G. Möller amerikai genetikus kísérletei a röntgensugárzás hatásairól a Drosophilán és a mutációk kvantitatív elszámolására szolgáló módszerek kifejlesztésében. G. Meller munkája számos kísérleti tanulmányt eredményezett különböző tárgyak röntgenfelvételein. Ennek eredményeként általános mutagén hatásuk jött létre. Később kiderült, hogy más típusú sugárzás, mint például az UV, valamint a magas hőmérséklet és néhány vegyi anyag is mutagén hatású. Az első kémiai mutagéneket a 30-as években fedezték fel. a Szovjetunióban V.V. Sakharova, M.E. Lobasheva és S.M. Gershenzon és alkalmazottai. Néhány évvel később ez a terület széles körű, különösen az A.I. Rapoport a Szovjetunióban és S. Auerbach Angliában.
A kísérleti mutagenezis területén végzett kutatás gyors fejlődést eredményezett a mutációs folyamat ismeretében, és számos, a gén struktúrájával kapcsolatos kérdés tisztázására.
A genetikai kutatás másik fontos területe a klasszikus genetika időszakában a genetikai folyamatok evolúcióban betöltött szerepének tanulmányozására irányult. Ezen a területen alapító munkája S. Wright, R. Fisher, J. Haldane és SS. Chetverikov. Munkáik révén megerősítették a darwinizmus alapelveinek érvényességét, és hozzájárultak egy új, modern szintetikus evolúciós elmélet megteremtéséhez, amely Darwin elméletének és populációs genetikájának szintézisének eredménye.
1940 óta a második periódus a világ genetikai fejlődésében kezdődött, amelyet molekulárisnak neveztek, a genetikai tudomány ezen területének vezető pozíciójának megfelelően. A molekuláris genetika gyors növekedésében a fő szerepe a biológusok szoros szövetsége volt a természettudományok más területein (fizika, matematika, kibernetika és kémia) dolgozó tudósokkal, amelyeken számos fontos felfedezés történt. Ebben az időszakban a tudósok megállapították a gén kémiai természetét, meghatározzák a működés és az ellenőrzés mechanizmusait, és számos fontosabb felfedezést tettek, amelyek a genetikát a modern természettudomány fejlődését meghatározó egyik fő biológiai tudományággá változtatták. A molekuláris genetika felfedezései nem gátolják, hanem csak azoknak a genetikai mintáknak a mögöttes mechanizmusait tárták fel, amelyeket a formális genetika fedezett fel.
J. Bidle és E. Tetum (USA) munkájával megállapították, hogy a Neurospora crassa kenyérformában mutációk blokkolják a celluláris metabolizmus különböző szakaszait. A szerzők azt javasolják, hogy a gének szabályozzák az enzim bioszintézisét. Megjelenett egy tézis: „egy gén - egy enzim”. 1944-ben az amerikai tudósok (O. Avery, C. McLeod és M. McCarthy) által végzett bakteriális genetikai transzformációról szóló tanulmány kimutatta, hogy a genetikai információ hordozója a DNS. Ezt a következtetést később megerősítettük a transzdukció jelenségének vizsgálatában (J. Lederberg és M. Zinder, 1952) - az információ bakteriális sejtről a másikra történő átvitelére fág DNS-sel.
Ezek a tanulmányok fokozott érdeklődést mutattak a DNS szerkezetének tanulmányozása iránt, ami 1953-ban létrehozta a DNS molekula modelljét J. Watson (amer. Biologist) és F. Crick (eng. Chemist). Dupla hélixnek hívták, mivel a modell szerint két spirálisan csavart polinukleotid láncból épült. A DNS olyan polimer, amelynek monomerei nukleotidok. Minden nukleotid dezoxiribóz-penta-szén-cukorból, foszforsav-maradékból és négy nitrogénbázisból (adenin, guanin, citozin és timin) áll. Ez a munka fontos szerepet játszott a genetika és a molekuláris biológia továbbfejlesztésében.
E modell alapján először feltételezték (F. Crick), majd kísérletileg bizonyították (M. Meselson és F. Stal, 1957) a DNS-szintézis félig konzervatív mechanizmusa, amelyben a DNS-molekula két különálló láncra oszlik, amelyek mindegyike szolgál mátrix a lánylánc szintéziséhez. A szintézis alapja az E. Chargaff (1945) által korábban definiált komplementaritás elve, amely szerint két DNS-lánc nitrogénbázisa párban egymással szemben van elrendezve, és az adenin csak a timinnel (A-T) és a guaninnal citozinnal (G-C) kapcsolódik. A modell létrehozásának egyik következménye a genetikai kód - a genetikai információ rögzítésének elve - megfejtése volt. A különböző országokban számos kutatócsoport dolgozott ezzel a problémával. Sikerünk Amer. Genetika M. Nirenberg (Nobel-díjas), akinek laboratóriumában dekódolták az első kódszót. Ez a szó a YYY triplett volt, amely három nukleotid szekvenciája azonos nitrogén bázissal - uracil. Az ilyen nukleotidok láncából álló mRNS molekula jelenlétében egy monoton fehérjét szintetizáltunk, amely ugyanazon aminosav, fenilalanin szekvenciálisan kapcsolt maradékait tartalmazza. A kód további dekódolása a technológia kérdése volt: a bázisok különböző kombinációival rendelkező mátrixokat használva a tudósok egy kódtáblázatot állítottak össze. A genetikai kód minden jellemzőjét azonosították: egyetemesség, triplett, degeneráció és nem átfedés. A genetikai kód dekódolását a tudomány és a gyakorlat fejlesztése szempontjából hasonlítják össze a fizika nukleáris energia felfedezésével.
A genetikai kód megfejtése és a genetikai információ rögzítésének elvének meghatározása után a tudósok azt gondolták, hogy az információ hogyan kerül át a DNS-ből a fehérjébe. Ennek a problémának a tanulmányozása a genetikai információ megvalósításának mechanizmusának teljes leírásával zárult le, amely két szakaszból áll: transzkripció és fordítás.
A gén kémiai természetének és hatásának meghatározása után felmerült a kérdés, hogy miként szabályozzák a géneket. Először a francia biokémikusok F. Jacob és J. Monod (1960) tanulmányaiban hallották, akik egy olyan rendszert fejlesztettek ki, amely szabályozza az E. coli sejtében laktóz-emésztés folyamatát szabályozó géncsoportot. A bakteriális operon fogalmát olyan komplexként mutatták be, amely egyesíti az összes gént (mind a strukturális, mind a szabályozó géneket), amely az anyagcsere bizonyos kapcsolatait szolgálja. Később a rendszereik helyességét kísérletileg igazolták az operon különböző szerkezeti egységeit érintő különböző mutációk tanulmányozásában.
Az eukarióta gének szabályozása fokozatosan fejlődött. Ezt megkönnyítette néhány gén egy folytonos szerkezetének kialakítása és a kötési mechanizmus leírása.
A gének szerkezetének és működésének tanulmányozása a 70-es évek elején történt előrehaladás hatása alatt. 20. század. A genetikusok azt gondolták, hogy manipulálják őket, először a sejtről a cellába történő áthelyezéssel. Így megjelent a genetikai kutatás új iránya - a géntechnológia.
Ennek az iránynak az alapja a kísérletek, amelyek során kidolgozták az egyes gének beszerzésére szolgáló módszereket. 1969-ben J. Beckwith laboratóriumában laktózoperont izoláltunk az Escherichia coli kromoszómájából a transzdukció jelenségével. 1970-ben G. Korán vezetése alatt álló kollektív volt az első, aki kémiai génszintézist végzett. 1973-ban kidolgoztunk egy módszert a DNS-fragmensek - gén-donorok - előállítására, a DNS-molekulát vágó restrikciós enzimek alkalmazásával. Végül pedig a fordított transzkripciós jelenség alapján géngyűjtési eljárást fedeztek fel, amelyet 1975-ben D. Baltimore és G. Temin fedezett fel. Az idegen gének plazmidokon, vírusokon, bakteriofágokon és transzpozonokon (mobil genetikai elemek) alapuló sejtekbe történő beviteléhez különböző vektorokat terveztünk - hordozó molekulákat, amelyek az átviteli folyamatot hajtották végre. A vektorral komplexet rekombináns molekulának nevezzük. Az első rekombináns fág alapú DNS molekulát 1974-ben tervezték meg (R. Murray és D. Murray). 1975-ben az integrált génekkel rendelkező sejtek és fágok klónozására szolgáló módszereket fejlesztettek ki.
Már a 70-es évek elején. A géntechnológia területén végzett kísérletek első eredményeit kaptuk. Így egy rekombináns molekulát inszertáltunk az E. coli sejtébe, amely két különböző antibiotikum rezisztencia gént tartalmaz (tetraciklin és sztreptomicin), majd a sejt mindkét gyógyszerre rezisztenciát szerez.
Fokozatosan bővült a vektorok és a bevezetett gének, és javult az átviteli technológia. Ez lehetővé tette az ipari célú géntechnológiai módszerek (biotechnológia) széles körű alkalmazását, elsősorban az orvostudomány és a mezőgazdaság érdekében. Terveztek baktériumokat - biológiailag aktív anyagok termelőit. Ez lehetővé tette az ilyen alapvető humán gyógyszerek - például inzulin, szomatosztatin, interferon, triptofán stb. - szintézisének megfelelő méretbeállítását. Ennek eredményeként nagyszámú transzgenikus növény keletkezett, amely értékes tulajdonságok tulajdonosai lettek (kártevőkkel szembeni ellenállás, aszály, magas fehérjetartalom stb.). idegen gének bejuttatása a genomba.
A 70-es években. a különböző objektumok genomjainak szekvenálásával kezdődött a munka, kezdve a bakteriofágokkal és véget ért az emberekkel.
Különös figyelmet kell fordítani a „Human Genome” nemzetközi genetikai programra, amelynek célja az emberi genetikai kód teljes dekódolása és a kromoszómák feltérképezése. A jövőben az orvosi genetika új területének - génterápia - intenzív fejlesztését tervezik, amely segít csökkenteni a káros gének megnyilvánulásának kockázatát, és ezáltal korlátozni a maximális genetikai terhelést.
A genetika fejlődésének története Oroszországban
A genetika megjelenése Oroszországban a huszadik század második évtizedében történt. Az első nemzeti genetikai iskola alapítója Jurij Alexandrovich Filipchenko volt. 1916-ban elkezdett előadást tartani a Szentpétervári Egyetemen „Az öröklés és az evolúció doktrínája” című kurzusról, amelyben Mendel törvényei és T. Morgan tanulmányai központi szerepet játszottak. A Morgan könyvének „Gene Theory” című könyvét engedélyezte. A Yu.A. tudományos érdekei. Filipchenko a minőségi és mennyiségi jellemzők öröklődésének és variabilitásának területén feküdt. Különös figyelmet fordított a változékonyság statisztikai törvényeire. YA Filipchenko számos kitűnő könyvet írt, köztük a „Genetika” tankönyvet, mely szerint a biológusok több generációja tanulmányozott hazánkban.
Ugyanebben az időszakban még két tudományos genetikai iskola jött létre: az egyik a Moszkvai Kísérleti Biológiai Intézetben, Nikolai Konstantinovich Koltsov vezetésével, a másik pedig Nikolai Ivanovich Vavilov vezetésével Saratovban jött létre, ahol egyetemi tanárgá választották, és végül Leningrádban lett. az All-Union Institute of Plant Industry (VIR) alapján.
NK Koltsov egy nagy kísérleti biológiai kutatóintézetet vezetett Moszkvában. Ő volt az első, aki kifejezte az öröklési hordozók makromolekuláris szervezésének (kromoszómák) és az önduplázódást, mint a genetikai információ továbbításának mechanizmusát. Ötletek N.K. Koltsov erős hatást gyakorolt \u200b\u200baz adott időszak híres tudósaira, nemcsak a biológusokra, hanem a fizikusokra is, akiknek a gén szerkezetének tanulmányozása a molekuláris genetika kialakulásához vezetett. N.K. tudományos iskolájából. Koltsov olyan nagy genetika, mint A.S. Serebrovsky, B.L. Astaurov, N.P. Dubinin, N.V. Timofeev-Resovsky, V.V. Szaharov és mások.
Kiváló genetikus és tenyésztő N.I. Vavilov széles körben elismerte munkáját a világ mezőgazdaságának és növényi erőforrásainak tanulmányozásában. Ő a szerzők a termesztett növények származási központjainak és sokféleségének elméletének, valamint az immunitás elméletének, valamint az örökletes változások homológ sorozatának törvénye. Emellett létrehozta a mezőgazdasági és műszaki üzemek világgyűjteményét, köztük a búzafajták híres gyűjteményét. NI Vavilovnak nemcsak a hazai, hanem a külföldi tudósok körében is nagy presztízs volt. A világ minden országából származó tudósok a Leningrádban létrehozott All-Union Institute of Plant Industry-ban (VIR) dolgoztak. Az érdemek elismerése N.I. Vavilovot az 1937-ben Edinburghban tartott Nemzetközi Genetikai Kongresszus elnökévé választották. A körülmények azonban nem tették lehetővé N.I. Vavilov részt vesz a kongresszuson.
Az elméleti genetika kialakulásának komoly hozzájárulását Alexander S. Serebrovsky, a moszkvai egyetem professzora és fiatal kollégái, az N.P. Dubinin, B.N. Sidorova, I.I. Agola és mások. 1929-ben felfedezték a Drosophilában bekövetkezett lépcsőzetes allélicizmust, ami az első lépés annak elhagyása felé, hogy a gén oszthatatlan volt a genetika között. A génszerkezet központi elmélete megfogalmazásra került, amely szerint a gén kisebb alegységekből áll, amelyek egymástól függetlenül mutálódhatnak. Ezek a tanulmányok ösztönzik a gén szerkezetét és működését vizsgáló tanulmányok kialakulását, ami a gén komplex belső szervezésének modern koncepciójának kialakulását eredményezte. Később (1966-ban) a mutációs elmélet területén végzett munkák ciklusához, N.P. Dubinin elnyerte a Lenin-díjat.
A 40-es évek elejére. 20. század. a Szovjetunióban a genetika virágzó állapotban volt. A fentieken kívül B.L. Astaurov a selyemhernyó szexuális szabályozásáról genetikai módszerekkel; G.A. Levitsky, A.A. Sapegina, K.K. Meister, A.R. Zhebraka, N.V. Tsitsina a genetikáról és a tenyésztésről; MF Ivanova a genetikáról és az állattenyésztésről; VV Sakharova, M.E. Lobasheva, S.M. Gershenzon, I.A. Rapoport a kémiai mutagenezisről; SG Levita és S.N. Davidenkova az emberi genetikáról és sok más tehetséges tudós munkájáról.
A szovjetunióban a kapitalista világgal való szembenézés politikai helyzete azonban a második világháború elején a genetika területén dolgozó tudósok üldözéséhez vezetett, melyet idealista burzsoá tudománynak nyilvánítottak, és a világ imperializmusának képviselőit. Az elnyomások sok híres tudós, köztük N.I. Vavilova, M.E. Lobasheva, GD Karpechenko, S.M. Gershenzon és sokan mások. A genetikát néhány évtizeddel ezelőtt leesették. A genetikai tudomány lebontásában jelentős szerepet játszott az ETC. Lysenko. Egyszerű agronómusként a génről alkotott absztrakt elképzeléseivel nem tudott felállni a klasszikus genetika szintjére, és ezért egyszerűen tagadta a Mendel törvényeit, a Morgan öröklődésének kromoszóma elméletét, a mutációk tanulmányozását. Lysenko tudományos következetlenségét a gazdálkodás gyors növekedésének nagylelkű ígéreteivel fedte fel, az általa támogatott növényi feldolgozás módszereivel, amelyek személyesen I.V. Sztálin. Lysenko pajzsként használta a kiemelkedő tenyésztő I.V. munkáját. Michurina. A világtudománytól eltérően genetikánk Michurin néven vált ismertté. Egy ilyen „becsület” azt a tényt eredményezte, hogy Michurin-t Lysenko, az ötletek csatlakozója, aki az utóbbi tevékenységeinek összeomlása után sem hagyta el a tudósot. Valójában I.V. Michurin kiemelkedő gyakorló tenyésztő, gyümölcstermelő volt, aki soha nem volt köze a genetikai tudomány elméleti alapjainak fejlesztéséhez.
A házi tudományt végül a 60-as évek közepéig „lizenkoizmust” tisztították. Sokan az elnyomástól szenvedő tudósok közül, akik sikerült túlélniük, beleértve az N.V. Timofeev-Resovsky, M.E. Lobashov, V.V. Szaharov és mások. A megőrzött hagyományok és a diákok által rejlő nagy potenciál hozzájárult a gyors előrelépéshez, bár a világszintű lemaradás természetesen érezte magát. Az orosz genetikusok új generációját azonban felvetették, akik ezt a tudományt az előző szintre vitték. És ismét a világhírű tudósok sorai az orosz nevekkel bővültek: A.N. Belozersky, V.A. Engelhardt, S.I. Alikhanyan, R. B. Khesin, A.S. Spirina, S.V. Shestakova, S.G. Inge-Vechtomova, Yu.P. Altukhov és sokan mások.
Azonban a tudományos munkatársak külföldre történő kiáramlásához vezető szerkezetátalakítás okozta új társadalmi megfordulások ismét megakadályozták tudományunkat a megfelelő státusz megszerzésében. Reméljük, hogy a fiatalabb nemzedék, az előző világítótestek alapjaira támaszkodva, képes lesz teljesíteni ezt a nemes küldetést.
Előadás terv
A genetika tárgya. Az öröklődés és a variabilitás jelenségének lényege.
A genetika módszerei.
A genetika fejlődésének rövid története.
Genetikai tanulmányok a Fehérorosz Köztársaságban
A genetika kapcsolata más tudományokkal.
A genetika értéke.
Kérdésre. A genetika tárgya. Az öröklődés és a variabilitás jelenségének lényege.
A genetika az élő szervezetek öröklődésének és variabilitásának tudománya, valamint a kontroll módszerei; ez az a tudomány, amely az élő szervezetek jellemzőinek öröklését és változékonyságát vizsgálja.
átöröklés- Ez az
1) a szervezetek képessége saját fajta előállítására;
2) a szervezetek azon képessége, hogy átadják (örököljék) sajátosságait és tulajdonságait generációról generációra;
3) a tulajdonságok változatosságának megőrzése a generációk változásakor.
változékonyság - az a szervezet képessége, hogy a test vagy annak egyes részei, valamint funkciói szerint változnak.
Variabilitás
1) különböző formájú jelek (változatok) megléte;
2) az egyének (testrészek vagy szervezetcsoportok) közötti különbségek kialakulása egyedi alapon.
A vonás örökségének fő típusai
Közvetlen öröklésahol a jelek változása nem változik a nemzedékről nemzedékre.
- a növények vegetatív reprodukcióján;
- a növények önszennyeződése során;
- a fajtatiszta állatok szaporodása és a fajtatiszta növények keresztezése során.
Közvetett öröklés - Ez a fajta örökség, amelyet az állatok szexuális szaporodása és a növények magszaporodása során figyeltek meg.
A közvetett örökség tanulmányozásához hibridizációra van szükség - a genotípusban eltérő szervezetek átkelése.
Közvetett örökségben az egyes generációkban a jellemzők bizonyos változatai jelennek meg (az ilyen jellemzőket dominánsnak, „dominánsnak” nevezik), és más variánsok átmenetileg „eltűnhetnek”, majd a következő generációkban megnyilvánulhatnak (az ilyen jellemzőket recesszívnek, „visszavonulásnak” nevezik).
A vonás örökségének összetett típusai nagyon nehéz előre jelezni a jelek új variációinak megjelenését. Bizonyos esetekben a "hirtelen" jelek új variációi vannak, amelyeket a szülők, nagyszülők vagy nagynénik nem találtak. A tulajdonságok komplex öröksége mind a környezeti feltételeknek a szervezet fejlődésére gyakorolt \u200b\u200bhatása, mind az új gének vagy a szervezetben jelen lévő gének új kombinációi következtében lehetséges.
Kérdésre. A genetika módszerei.
A genetikában, mint más tudományok, a kutatásban számos módszert alkalmaznak. A genetika saját kutatási módszerekkel rendelkezik:
Hibridológiai elemzés - a fő módszer, amelyben a szülők céltudatos keresztezését hajtják végre bizonyos jellemzőkkel, és megfigyelik e tulajdonságok megnyilvánulását az utódok generációiban.
A hibridológiai elemzés alapelvei:
1. Kezdeti egyénekként (szülők) történő felhasználás, olyan formák, amelyek keresztezéskor nem oszlanak meg, azaz állandó formák.
2. Az alternatív jellemzők párjainak örökségének elemzése, azaz a két egymást kizáró lehetőség által képviselt jellemzők.
3. Az egymást követő kereszteződések során menekülő formák mennyiségi leírása és matematikai módszerek alkalmazása az eredmények feldolgozásában.
4. Az utódok egyéni elemzése minden szülőtől.
5. Az átkelés eredményei alapján összeállítják és elemzik az átkelés mintáját.
leszármazási - a törzskönyvek elemzése, és lehetővé teszi az öröklési (domináns, recesszív, autoszomális vagy nemi összefüggésű) tulajdonságok meghatározását, valamint annak monogenitását vagy polioxicitását. A kapott információk alapján megjósolható a vizsgált tulajdonság megjelenésének valószínűsége az utódokban, ami nagyon fontos az örökletes betegségek megelőzésében.
citogenetikai - a kromoszómák vizsgálata: számuk számozása, a szerkezet leírása, a sejtfelosztás során fellépő viselkedés, valamint a kromoszóma szerkezet változásainak összefüggése a karakterek variabilitásával.
biokémiai - az enzimrendszerek aktivitásának tanulmányozása alapján. Az aktivitást vagy maga az enzim aktivitása, vagy a végső reakciótermékek mennyisége határozza meg, amelyeket ez az enzim szabályoz. Az enzimrendszerek aktivitásának vizsgálata lehetővé teszi az anyagcsere-betegségek okát okozó génmutációk azonosítását, például a fenilketonuria, a sarlósejtes anaemia.
molekuláris - lehetővé teszi a DNS-fragmensek elemzését, az egyes gének megtalálását és izolálását, a nukleotidok szekvenciájának megállapítását (örökletes információt hordoz).
kérdés. A genetika fejlődésének rövid története.
Az ókori Görögország híres orvosa, Hippokratész úgy gondolta, hogy a tojássejtben vagy az anya testében egy kicsi, de teljesen kialakult, előformázott szervezetnek kell lennie. Ezek a meggyőződések később előformizmusnak (latin nyelvűek. Preforraatio - pre-formáció) váltak ismertté. Az előformisták közötti vita csak arról szólt, hogy hol található ez a szervezet - a nőies vagy férfias.
Ellentétes nézetek, amelyek szerint a szervezet strukturált, homogén tömegből fejlődik ki, amelyet először Arisztotelész kifejezett, később kifejlesztette az epigenesis nevét (a görög Epi-től és a genesis-fejlődéstől).
Charles Darwin először tudományos alapokra helyezte a biológiát. Megmutatta, hogy az evolúció és a kiválasztás alapja az öröklés, a variabilitás és a kiválasztás hatása. Ezek a rendelkezések a genetika minden későbbi fejlesztésének alapját képezték.
Első szakasz a tudomány fejlesztése.
Jellemzője, hogy G. Mendel (1865) felfedezte az örökletes tényezők diszkrétitását (oszthatóságát) és egy hibridológiai módszer kifejlesztését, az öröklődés tanulmányozását, vagyis a szervezetek keresztezésének szabályait, valamint az utódokban lévő jelek figyelembevételét.
G. Mendel felfedezéseinek jelentőségét azután állapították meg, hogy 1900-ban ismét három biológus egymástól újra felfedezte a törvényeit: de Vries Hollandiában, K. Correns Németországban és E. Cermak Ausztriában.
1901-1903 Hugo de Vriesmutációs elméletet mutattak be, amely nagy szerepet játszott a genetika továbbfejlesztésében.
A dán botanikus munkája fontos volt. Wilhelm Ludwig Johansen, aki tanulmányozta az örökség mintáit a tiszta babsorokon. Szintén megfogalmazta a „népesség” fogalmát (egy korlátozott területen élő és tenyésztő ugyanazon faj élőlénycsoportja), azt javasolta, hogy a „gén” fogalmát a „genotípus” és a „fenotípus” fogalmának meghatározására hívják fel.
Második szakasz
Jellemzője az öröklődési jelenségek tanulmányozására való átmenet a sejtek szintjén (citogenetika). T. Boveri (1902-1907), W. Satton és E. Wilson (1902-1907) megállapították a mendeli öröklési törvények és a kromoszómák eloszlásának összefüggését a sejtosztódás folyamatában (mitózis) és a csírasejtek érésében (meiosis).
Az öröklődés kromoszóma elméletének megalapozásában döntő jelentőségű volt az T. G. Morgan amerikai amerikai genetikus és munkatársai (1910–1911) által végzett legyek.
Morgan szintén létrehozta a nemhez kötődő vonások örökségét.
A következő lépés a kromoszóma-gének kémiai természetének meghatározása volt. Szovjet genetikus N.K. Koltsov volt az egyik első, aki ötletet készített a makromolekuláris természetükről (1927) és N.V. Timofeev-Resovsky és társszerzők az 1930-as évek közepén. 20 in. kiszámította a gén hozzávetőleges mennyiségét.
1925-ben először szovjet mikrobiológusok G.A. Nadson és G.S. Filippov kimutatta, hogy az élesztősejtek ionizáló sugárzással történő besugárzása után különböző rádió sugárzások keletkeznek, amelyek tulajdonságai az utódokban reprodukálódnak. 1927-ben, Möller (N.J. Muller) a gyümölcsök legyőzésével végzett pontos kísérletekben, figyelembe véve a sugárzási dózist, megállapította az új örökletes mutációk megjelenését. Később I.A. Rapoport és Auerbach (Ch. Auerbach) felfedezték a mutagenezis jelenségét a vegyi anyagok hatására.
Harmadik szakasz
A molekuláris biológia eredményeit tükrözi, és a pontos tudományok - a fizika, a kémia, a matematika, a biofizika stb. - módszereinek és elveinek alkalmazásával kapcsolódik az életjelenségek molekuláris szintű tanulmányozásához. A genetikai kutatások tárgyai gombák, baktériumok, vírusok lettek.
Ebben a szakaszban tanulmányozták a gének és az enzimek közötti kapcsolatot, és az „egy gén - egy enzim” elméletet fogalmazták meg (J. Bidle és E. Tatem, J. Lederberg, 1940): minden gén egy enzim szintézisét szabályozza; Az enzim viszont egy, a szervezet külső vagy belső tulajdonságának megnyilvánulását alkotó biokémiai átalakulások egy reakcióját szabályozza.
1953-ban F. Crick és J. Watson, a genetikusok és a biokémikusok kísérleteinek eredményei alapján, a röntgenszerkezeti elemzés adatai alapján kettős hélix formájában létrehozott DNS szerkezeti modelljét. Az általuk javasolt DNS-modell jó egyezést mutat ennek a vegyületnek a biológiai funkciójával: a genetikai anyag önmásolásának képességével és a generációtól a sejtektől a sejtekig.
Az elmúlt évtizedben a molekuláris genetika új iránya - géntechnológia - olyan technológiai rendszer, amely lehetővé teszi a biológus számára, hogy mesterséges genetikai rendszereket állítson elő.
Orosz Föderáció Oktatási Szövetsége
Állami felsőoktatási felsőoktatási intézmény
"Dél-Urál Állami Egyetem"
Közgazdaságtudományi Kar
Közgazdasági, menedzsment és befektetési osztály
A genetika története. Az orosz tudósok hozzájárulása
KIVONAT
a "tudomány fogalmai" fegyelemről
kockás
OM Baev
hallgatói csoport EiU-232
AI Kuleshov
________________________ 2010
Az absztrakt védett
értékeléssel
_____________________________
________________________ 2010
BEVEZETÉS
A genetika - az öröklés és változékonyság tudománya - a 20. század elején alakult ki, miután a kutatók figyelmet szenteltek az 1865-ben felfedezett G. Mendel törvényeinek, de 35 évig felügyelet nélkül maradt. Rövid idő alatt a genetika kiterjedt biológiai tudománygá fejlődött, számos kísérleti módszerrel és irányokkal. A genetika nevét William Batson angol tudós javasolta 1906-ban. A genetika fejlődésének klasszikus időszakának kutatói felfedezték az öröklés főbb törvényeit, és bebizonyították, hogy az örökletes tényezők (gének) a kromoszómákban koncentrálódnak. A genetikai információk tárolásának és értékesítésének mintáinak tanulmányozásában további előrelépést tartottak vissza két okból. Először is, a gének mélyebb vizsgálatához kapcsolódó túl kiterjedt kísérletek miatt, másrészt a gének munkájának megértésének lehetetlensége nélkül, a genetikai folyamatokban részt vevő molekulák átalakulásának alapos tanulmányozása nélkül. A mikroorganizmusok genetikai vizsgálatára való áttérés, amely lehetővé tette a sok nehézség elkerülését, egészen természetes volt. Ez az átmenet az 50-es években történt. 1941-ben J. Beadle és E. Tatum egy rövid cikket tett közzé: "A biokémiai reakciók genetikai ellenőrzése Neurospora ", amely jelentette az első genetikai kísérleteket a mikroorganizmusokkal.
Az utóbbi években ezek a tanulmányok széles körű alkalmazást kaptak, és számos biológiai objektumon végzik őket.
A dolgozat célja, hogy tükrözze az orosz tudósok által a genetika területén tett legfontosabb felfedezéseket, azok elemzését és jelentőségének meghatározását a tudomány számára.
A téma nyilvánosságra hozatalához mind a tudományos munkákat, mind a modern internetes erőforrásokat vették igénybe, amelyeknek megbízható adatokat és modern szempontokat kell biztosítaniuk.
1 GENETIKAI FEJLESZTÉS Oroszországban
A XVIII. Században a növények hibridizációjára irányuló kísérleteken kívül a XX. Század elején megkezdődtek az oroszországi genetika első munkái. mind a kísérleti mezőgazdasági állomásokon, mind az egyetemi biológusoknál, főként a kísérleti botanikában és az állattan.
Az 1917-1922-es forradalom és polgárháború után. a tudomány gyors szervezeti fejlődését kezdte. Az 1930-as évek végére a Szovjetunióban létrehozták a kutatóintézetek és kísérleti állomások kiterjedt hálózatát (mind a Szovjetunió Tudományos Akadémiáján, mind a Lenin All-Union Academy of Agricultural Sciences (VASKhNIL)), valamint a genetikai egyetemek. A tendencia elismert vezetői N. Vavilov N., Koltsov N., S. Serebrovsky, S. S. Chetverikov és mások, a Szovjetunióban külföldi genetikusok, köztük T. Khgan, G. Moeller fordításai kerültek kiadásra. Számos genetikus vett részt nemzetközi tudományos csereprogramokban. Az amerikai genetikus G. Möller dolgozott a Szovjetunióban (1934-1937), a szovjet genetikusok külföldön dolgoztak. N. V. Timofeev-Resovsky - Németországban (1925 óta), FG Dobzhansky - az USA-ban (1927 óta).
Az 1930-as években T. Lysenko és I. I. Prezent energetikai tevékenységeihez kapcsolódó genetikusok és tenyésztők soraiban megoszlott. A genetikusok kezdeményezésére egy sor vitát tartottak (a legnagyobb 1936-ban és 1939-ben), amelynek célja a Lysenko-megközelítés elleni küzdelem.
Az 1930-1940-es évek fordulóján. az úgynevezett Nagy Terror alatt a CPSU (b) központi bizottságának személyzete, aki a genetikát felügyelte, és számos kiemelkedő genetikus letartóztatták, sokan lőttek vagy meghaltak börtönökben (ideértve N. I. Vavilovot is). A háború után a vita új erővel folytatódott. A genetikai szakemberek a nemzetközi tudományos közösség jogosultságára támaszkodva ismét megpróbálták a mérlegeket javítani, de a hidegháború elején a helyzet jelentősen megváltozott. 1948-ban, az Agrártudományi Akadémia augusztusi ülésén, T. D. Lysenko, Sztálin József támogatásával, hamis tudományt hirdetett meg. Lysenko kihasználta a tudomány pártvezetésének inkompetenciáját, "az ígéretet a pártnak" az új, rendkívül produktív gabonafajták ("elágazó búza") gyors létrehozását, stb. Ebből a ponttól kezdve kezdődött a genetika üldöztetésének ideje, amely Lysenkovism néven vált ismertté, és addig folytatódott, amíg N. Hruštšovot eltávolították. a CPSU Központi Bizottságának főtitkára 1964-ben
Személy szerint T. D. Lysenko és támogatói átvették a Szovjetunió Tudományos Akadémia Biológiai Intézetének, a Mezőgazdasági Tudományos Akadémia és az egyetemi tanszékek intézményeit. Megjelentek az új iskolák és egyetemek tankönyvei, amelyeket a "Michurinsk Biológia" álláspontjából írtak. A genetika kénytelen volt elhagyni tudományos tevékenységét, vagy radikálisan megváltoztatni a munka profilját. Néhányan képesek voltak folytatni a genetika kutatását a sugárzás és a kémiai veszélyek tanulmányozására irányuló programok keretében a TD Lysenko és támogatói által ellenőrzött szervezeteken kívül.
A DNS szerkezetének, a gének fizikai bázisa (1953) felfedezése és dekódolása után a genetika helyreállítása az 1960-as évek közepén kezdődött. Az RSFSR oktatási minisztere, VN Stoletov, széles körű vitát kezdeményezett a lizenkoisták és a genetikusok között, és ennek eredményeként számos új genetikai munkát jelentettek meg. 1963-ban kiadták M. E. Lobashev „Genetika” egyetemi tankönyvét, amely később több publikáción ment keresztül. Hamarosan megjelent egy új iskolai tankönyv, Általános Biológia, amit Yu I. Polyansky szerkesztett, másokkal együtt, a mai napig.
Következtetési szakasz első
Az oroszországi genetika fejlődése nehéz módon zajlott, a hatóságok üldöztetése miatt, ami jelentősen lelassította e tudomány fejlődését.
2 NIKOLAY IVANOVICH VAVILOV ÉS HATÁROZATA GENETIKÁHOZ
Nikolai Ivanovics Vavilov (november 13 (25), 1887, Moszkva, orosz birodalom - 1943. január 26., Saratov, RSFSR, Szovjetunió) —Az orosz és szovjet genetikai tudós, botanikus, tenyésztő, földrajz, akadémikus a Szovjetunió Tudományos Akadémiájának az ukrán SSR-ben és a Mezőgazdasági Tudományos Akadémián. Elnök (1929-1935), az Agrártudományi Akadémia alelnöke (1935-1940), az All-Union Geográfiai Társaság elnöke (1931-1940), alapító (1920) és állandó letartóztatásig, az Európai Unió Növényipari Intézetének igazgatója (1930-1940), a Genetikai Intézet igazgatója Szovjetunió (1930-1940), a Szovjetunió Tudományos Akadémia Expedíciós Bizottságának tagja, a Szovjetunió Népi Bizottságának igazgatótanácsának tagja, a keleti tanulmányok szövetségének elnökségének tagja. 1926-1935-ben tagja volt a Szovjetunió Központi Végrehajtó Bizottságának, 1927-1929-ben az All-Russian Központi Végrehajtó Bizottság tagja volt.
A botanikus és agronómiai expedíciók szervezője és résztvevője, amely a kontinensek legnagyobb részét (Ausztrália és Antarktisz kivételével) fedte le, mely során felfedezte a termesztett növények formálásának ősi gyökereit. Létrehozta a termesztett növények világközpontjainak doktrínáját. Megalapozta a növényi immunitás doktrínáját, felfedezte a homológ sorozatok törvényét a szervezetek örökletes változékonyságában. Jelentősen hozzájárult a biológiai fajok elméletének fejlesztéséhez. Vavilov vezetésével a világ legnagyobb termesztett növényi magvak gyűjteménye jött létre. Megalapozta a terepi növényfajták állami vizsgálati rendszerét. Megfogalmazta az ország fő tudományos központjának a mezőgazdasági tudományok tevékenységének alapelveit, létrehozta a tudományos intézmények hálózatát ezen a területen.
2.1 A növényi immunitás doktrínája
Vavilov a növényi immunitást strukturális (mechanikai) és vegyi anyagokra osztotta. A növények mechanikai immunitása a gazdanövény morfológiai jellemzői miatt, különösen a védőeszközök jelenléte, amelyek megakadályozzák a kórokozók bejutását a növények testébe. A kémiai immunitás a növények kémiai tulajdonságaitól függ.
2.2 Az örökletes variabilitás homológ sorozata
Az „Örökletes variabilitás homológ sorozatának törvénye” című munkában Vavilov bemutatta az „örökletes változékonyság homológ sorozatának” fogalmát. A koncepciót az örökletes variabilitás jelenségeiben a paralelízisek vizsgálatában vezették be a szerves vegyületek homológ sorozatával analóg módon.
A jelenség lényege, hogy a genetikai variabilitás vizsgálatakor a növények szoros csoportjaiban hasonló allélformákat találtak, amelyek különböző fajokban ismétlődtek (például gabona szalma csomók, antocianin színezéssel vagy anélkül, füle tüskékkel vagy anélkül stb.) ). Az ilyen ismételhetőség jelenléte lehetővé tette a még nem észlelt allélok előrejelzését, amelyek a szelekciós munka szempontjából fontosak. Az ilyen allélokkal rendelkező növények keresését a termesztett növények állítólagos származási központjaira irányuló expedíciókon végeztük. Emlékeztetni kell arra, hogy ezekben az években a mutagenezis mesterséges indukciója a vegyi anyagokkal vagy az ionizáló sugárzásnak való kitettséggel még nem volt ismert, és a szükséges alléleket természetes populációkban kellett elvégezni.
A törvény első (1920) szövege két szabályszerűséget tartalmazott:
Az első minta, amely a szemet részletesen megvizsgálja az azonos nemzetségbe tartozó növények bármelyik vonalának formáinak tanulmányozása során, a közeli genetikai linoneonok faját és faját jellemző morfológiai és élettani tulajdonságok sorozata, a specifikus genotípusos variabilitás sorának párhuzamossága. Minél közelebb van a faj genetikailag, annál élesebb és pontosabb a morfológiai és fiziológiai karakterek sorozata.
A polimorfizmus második szabályossága, amely lényegében az elsőből következik, az, hogy nem csak a genetikailag kapcsolódó fajok, hanem a nemzetségek is azonosítanak identitásokat a genotípusos variációk sorozatában.
1923-ban Vavilov „A közelmúltbeli előrelépések a szelekció elméletében” című munkájában vita tárgyalt a törvényről, amelyben megmutatta, hogy a fajok és nemzetségek fajtájú különbségeinek megnyilvánulási formája miatt „feltétlenül előre látható és megfelelő formák találhatók meg a vizsgált növényben”. Valóban, a homológ sorozat törvénye alapján Vavilov és munkatársai több százszor előre jelezték bizonyos formák létezését, majd felfedezték őket. Vavilov megjegyezte, hogy „a variabilitás közös sorozata néha nagyon távoli, genetikailag független családokra jellemző”. Vavilov elismerte, hogy a párhuzamos variabilitás sorozata nem feltétlenül lesz teljes, és a természetes szelekció, a gének halálos kombinációi és a fajok kihalása következtében bizonyos kapcsolatoktól megfosztják. Azonban "annak ellenére, hogy a természetes szelekció és a sok kapcsolat kihalása hatalmas szerepet tölt be, ... nem nehéz nyomon követni az egymással összefüggő örökletes variabilitás hasonlóságait."
Bár a törvényt fenotípusos variabilitás tanulmányozása eredményeként fedezték fel, Vavilov kiterjesztette hatását a genotípusos variabilitásra: „A fajok fenotípusos variabilitásának ugyanazon nemzetségben vagy közeli nemzetségekben tapasztalható hasonlósága alapján, az evolúciós folyamat egysége miatt feltételezhetjük, hogy sokan közös gének a fajok és nemzetségek sajátosságai mellett. "
Vavilov úgy vélte, hogy a törvény tisztességes, nemcsak a morfológiai karakterek vonatkozásában, feltételezve, hogy a már létrehozott sorozat nem csak a hiányzó kapcsolatokkal bővül a megfelelő sejtekben, hanem fejlődik is, különösen a fiziológiai, anatómiai és biokémiai jellemzők tekintetében. Vavilov különösen megjegyezte, hogy a közeli növényfajokat a „kémiai összetétel hasonlósága, a közeli vagy azonos vegyi vegyületek előállítása” jellemzi. Amint azt Vavilov is kimutatta, a kémiai összetétel (például az illóolajok és az alkaloidok) intraspecifikus variabilitása főként állandó minőségi összetételű kvantitatív arányokra vonatkozik, míg a nemzetségen belül egyes fajok kémiai összetétele mennyiségi és minőségi szempontból is különbözik. Ugyanakkor a nemzetségen belül az „egyes fajokat általában elméletileg megtervezett kémikusok izomerjei vagy származékai jellemzik, és általában kölcsönös átmenetekkel kapcsolódnak egymáshoz”. A variabilitás párhuzamossága olyan szorosan jellemzi a szoros születést, hogy „alkalmasak a megfelelő kémiai komponensek keresésére”, valamint „arra, hogy szintetikusan szerezzenek be egy adott nemzetségben egy bizonyos minőségű vegyi anyagok keresztezésével”.
Vavilov rájött, hogy a törvény nemcsak a kapcsolódó csoportokban nyilvánul meg; a variabilitás párhuzamosságát „különböző családokban találták meg, nem genetikailag összefüggő, még a különböző osztályokban”, de a távoli családokban a párhuzamosság nem mindig homológ. "Hasonló szervek és ezek hasonlósága ebben az esetben nem homológ, csak analóg."
A homológ sorozat törvénye nem távolította el az összes nehézséget, mivel nyilvánvaló volt, hogy a fenotípusos tulajdonságok azonos változásai különböző géneknek tudhatók be, és az ezekben az években létező tudás szintje nem engedte közvetlenül a tulajdonságot egy adott génhez. A fajok és nemzetségek tekintetében Vavilov megjegyezte, hogy „még mindig nem génekkel foglalkozunk, amelyekről nagyon keveset tudunk, de bizonyos környezetben jelei vannak”, és ennek alapján inkább homológ karakterekről beszélt. "A távoli családok párhuzamossága esetén az osztályok természetesen nem is beszélhetnek azonos génekről a hasonló külső jelek esetében sem."
Annak ellenére, hogy a törvény eredetileg túlnyomórészt tenyésztett növények tanulmányozásán alapult, később, figyelembe véve a gombák, algák és állatok változatosságának jelenségét, Vavilov arra a következtetésre jutott, hogy a törvény univerzális jellegű, és nemcsak magasabb, hanem alacsonyabb szinten is nyilvánul meg. növényeket, valamint az állatokat.
A genetika fejlődésének jelentős hatása volt a törvény megfogalmazásának továbbfejlesztésére. Vavilov 1936-ban túlságosan kategorikusan hívta az első megfogalmazást: „Ez volt a genetika állapota abban az időben ...”. Szokás volt azt gondolni, hogy a „gének azonosak a közeli fajokban”, a biológusok „a gént stabilabban képviselték, mint jelenleg. Később megállapították, hogy a "közeli fajok hasonló külső jelek jelenlétében számos különböző gén jellemezhető." Vavilov megjegyezte, hogy 1920-ban "kevés figyelmet fordított a kiválasztás szerepére", a variabilitás mintáira összpontosítva. Ez a megjegyzés nem jelenti azt, hogy elfelejtettük az evolúció elméletét, mivel - ahogy maga Vavilov is hangsúlyozta - már 1920-ban a törvénye „mindenekelőtt az evolúciós tanításon alapuló pontos tények képletét képviselte”.
Vavilov úgy vélte, hogy az általa megfogalmazott törvény hozzájárult az akkori népszerűséghez az evolúciós folyamat alapját képező variabilitás természetes természetére (például L. S. Berg nomogeneziselméletére). Úgy vélte, hogy a különböző csoportokban rendszeresen ismétlődő örökletes variációk az evolúciós párhuzamok és a mimikriás jelenség alapjául szolgálnak.
Második fejezet Következtetés
Oroszország egyik első genetikusa, Vavilov jelentősen hozzájárult ennek a tudománynak a fejlődéséhez, megindítva a növények genetikai természetének tanulmányozását.
3 NIKOLAY KONSTANTINOVICH KOLTSOV
Nikolai Konstantinovich Koltsov (július 3 (15), 1872, Moszkva - 1940. december 2., Leningrad) - kiemelkedő orosz biológus, a mátrixszintézis ötletének szerzője.
A leíró biológia és a kísérleti biológia első lépéseiben végzett munkáját követően Koltsov finoman érezte a biológia fejlődésének trendjeit, és hamarabb felismerte a kísérleti módszer jelentőségét. A biológia minden területén kísérleti megközelítés iránti igényt prédikált, és az evolúciós tanulmányokban is megjósolta (a kísérleti módszereket leíró módszerekkel ellentétes). Nem egy egyszerű biológiai kísérletről volt szó, hanem a fizika és a kémia módszereinek használatáról. Koltsov többször is hangsúlyozta, hogy a radioaktív anyagok használatáról a sugárzó energia új formáinak, különösen a röntgensugaraknak és a kozmikus sugaraknak a felfedezésének biológia számára rendkívül fontos szerepe van. A szervezet egészének tanulmányozásához minden modern tudást a fizikai és a kolloid kémia területén kell felhasználni, a sejten belüli monomolekuláris rétegeket és az anyagok különböző átalakulásaiban betöltött szerepét tanulmányozni kell. "A biológusok arra várnak, hogy ezek a módszerek (röntgendiffrakciós elemzés) olyan fejlettek legyenek, hogy segítségükkel lehetővé válik az intracelluláris csontváz, szilárd fehérjék és más természetű struktúrák kristályszerkezetének tanulmányozása." Ez a gondolat prófétikus volt, és valójában megvalósult a DNS-molekula szerkezetének röntgenszerkezeti elemzésével. Koltsov másik ötlete prófétikusnak bizonyult, amelyben a biológiából a kémiaig is elment. Az a gondolat, hogy minden komplex biológiai molekula egy már létező hasonló molekulából származik, azt jósolta, hogy a vegyészek követik az új molekulák létrehozásának útját az összetett molekulák szükséges alkotórészeit tartalmazó oldatokban azáltal, hogy behozzák nekik az azonos szerkezetű kész molekulák magjait. . Azt írta: "Úgy gondolom, hogy csak így tudjuk szintetizálni a fehérjéket in vitro, de mégsem, de bizonyos, de bizonyos szintéziseket előre terveztek."
A látóterületen N.Koltsova állandóan genetikai kérdés volt. 1921-ben kísérleti munkát tett közzé: „A tengerimalacok festésének genetikai elemzése”. Vezetett genetikai vizsgálatok a gyümölcs legyek. Ezekben a munkákban a tudós a genetika és az evolúciós elmélet közötti legfontosabb kapcsolat létrehozását látta. Később megkezdődött a kémiai mutagenezis.
N.Koltsov mélyen megértette a genetika fontosságát az állattenyésztés gyakorlatában. 1918-ban megszervezte az Anikovskaya génállomást, amely a haszonállatok genetikájára specializálódott. Valamivel később egy másik baromfi üzemet szerveztek a Tula régióban. 1920 elején mindkét állomás egybeolvadt. 1925-ben az állomás megkapta a mezőgazdasági állatok genetikai központi állomásának nevét, amelynek igazgatója különböző években Koltsov és tanítványai voltak.
Következtetés 3. szakasz
NK Koltsov előre látta a genetika további fejlődését, így utat mutatva a követői felé. Hála neki és ötleteinek, hogy sok felfedezés történt, mint például a kémiai mutagenezis. Koltsov másik érdeme, hogy sok tehetséges embert vonzott az állomáson, akik később híressé váltak a teljes genetika alkotói és bizonyos típusú mezőgazdasági állatok kiválasztása.
4 A MUTÁCIÓK ARTIFICIÁLIS BEÁLLÍTÁSA
A kísérleti genetika legnagyobb eredménye az volt, hogy felfedezték, hogy különböző fizikai és kémiai ágensek segítségével mesterségesen mutációkat okozhat. Az orosz tudósok jelentősen hozzájárultak e lehetőség felfedezéséhez.
4.1 G.A hozzájárulása Nadson és tanítványai
Az egyik első kísérlet ezen a területen az orosz tudósok, Georgii Adamovich Nadson és fiatal munkatársa, Grigori Semenovich Filippov voltak, akik a Radiológiai és Radiológiai Intézetben dolgoztak.
Az élesztőben és az alsó gombákban mutációkat kapott rádium és röntgensugárzás hatására. A kísérletek eredményeképpen a telepek nagysága, alakja, színe eltérőek, az élesztősejtek megváltoztatták biokémiai tulajdonságaikat. Így felfedezték a sugárzás örökletes változások kiváltására való képességét.
Szintén ga Nadson E.Ya-val. Rokhlin közzétett egy sor olyan dokumentumot, amelyben először felvetették a kísérleti úton előállított mutánsok gyakorlati alkalmazásának kérdését.
Az élesztők és az alsó gombák mellett a GA Nadson laboratóriumában vizsgálták a sugárzás genetikai hatását a baktériumokra. Új irányt helyeztek a tudományba - a mikroorganizmusok populációs genetikája. A GA Nadson nagy figyelmet fordított a belső és külső tényezők szerepének tanulmányozására és elemzésére a kísérleti mutagenezis folyamatában, például, hogy a különböző típusú mikroorganizmusok hogyan reagálnak másképp a sugárzásra, és hogy a sugárzás intenzitása és dózisa hogyan befolyásolja a mutációk indukcióját. A GA Nadson iskolájának másik iránya a kémiai mutagenezis. 1928-ban tanítványai 1939-ben adatokat kaptak az élesztőben az örökletes változások előfordulásáról, a kőszénkátrány és a kálium-cianid hatására.
4.2. Timofeev-
Az 1930-as évek közepén egy elméletet fogalmaztak meg, amely leírja az ionizáló sugárzás aktiváló és mutagén hatásainak - az ún. „Célelmélet” - kinetikai függőségeit. Az elmélet alapjául szolgáló legfontosabb kísérleteket az 1931-1937 közötti időszakban végezték el. több kutató, köztük Nikolai Vladimirovich Timofeev-Resovsky, aki az ionizáló sugárzás kvantitatív biofizikájának egyik alapítója lett.
Timofeev-Resovskii kifejlesztette N. Koltsov ötleteit, aki azt javasolta, hogy a mátrixszintézis segítségével molekuláris örökletes szerkezeteket alakítsanak ki. Kutatást végzett a mutációs folyamat biofizikai elemzéséről, amely ezt követően molekuláris biológia kialakulásához vezetett, mint új szintetikus fegyelem. Timofeev-Resovskiy kimutatta, hogy a mutációs változások a kromoszómában lévő atomok viszonylag korlátozott csoportját érintik. Ez a felfedezés először a megértés molekuláris szintjéhez vezetett.
Nikolai Vladimirovics a radiobiológia egyik alapítója. Meg tudta állapítani, hogyan befolyásolja a sugárzás dózisa a mutációs folyamat intenzitását. Felfedezte az alacsony dózisú rádióstimuláció jelenségét és elemezte a sugárzás hatására a mutációk előfordulásának elsődleges kiváltó mechanizmusait.
Ez a kutató elsőként rámutatott arra, hogy az ionizáló sugárzásnak való kitettség közvetlen hatásai mellett (azaz rosszindulatú daganatok, égések, sugárbetegségek) súlyos káros mutációk és azok felhalmozódásának kockázata áll fenn a populációkban.
A mutációs folyamat kvantitatív elméletének egyik legfontosabb összetevője az orosz tudós tanulmányozása a közvetlen és fordított mutációk előfordulásának valószínűségéről.
1934-ben Timofeev-Resovsky egy sor ragyogó kísérletet hajtott végre, amely először megmutatta, hogy több recesszív mutáció kombinációja, amelyek mindegyike csökkenti az életképességet külön-külön, az ilyen kombinációkat hordozó egyének életképességének növekedéséhez vezethet. Ezek a tanulmányok lehetővé tették, hogy teljes mértékben megértsék a recesszió és a dominancia jelenségeinek evolúciós jelentőségét.
M. Delbrück (később Nobel-díjas), Nikolai Vladimirovich Timofeev-Resovsky együtt dolgozott a gének szerkezetének modellezésében. Ugyanebben az időszakban R. Rompu fizikussal együttműködve felfedezte és leírta a biológia „erősítő elvét”, amely a modern elméleti biológia egyik legfontosabb elve lett. Ennek az elvnek megfelelően az egyetlen változás megváltoztathatja az egész egyén tulajdonságait, és aktiválhatja azokat az erőket, amelyek több nagyságrenddel nagyobbak az elhasznált energia tekintetében.
4.3 Kémiai mutagenezis
Az első, aki felhívta a figyelmet a vegyi anyagok okozta mutációkra, N.K. Koltsov, aki javasolta a követői, V.V. Szaharovnak, hogy folytassa a kutatást ezen a területen.
VV Szaharov, a Koltsov tervén dolgozik, 10% -os jódoldatot használt. A padlóhoz kapcsolt látható mutációk elemzésével a második generáció utódaiban örökölt új mutációkat talált bemetszés és „Szilva” .
Első alkalommal a világon 1938-ban. V.V. Szaharov megfogalmazta a "mutációs faktorok specifikus hatásának" elképzelését, amely a spontán eredetű és a fizikai és kémiai mutagének által indukált mutációk természetében mutatkozott meg.
A mi időnkben V.V. Szaharov álláspontja, miszerint a mutagenezis sajátosságai egyrészt a befolyásoló tényező szerkezetéből, mind a szervezet jellemzőiből fakadnak, nem vesztette el jelentőségét, és a genetika egyik legfontosabb általánosítása. A mutációs folyamatról szóló tanulmánysorozat a belső tényezők (öregedés, beltenyésztés és hibridizáció) szerepének felfedezéséhez vezetett.
Az egyik első V.V. Szaharov felvetette a kolchicin és más mutagének mutáció utáni hatását a növényekre, amelyek ma nyertek értéket, és a biológiailag aktív anyagok kis adagjainak környezetvédőinek és kutatóinak elsődleges forrásává váltak.
Egy másik kutató, M.Ye Lobashev volt az első, aki a mutáció folyamatát a sérült genetikai struktúrák helyreállításával kapcsolta össze.
Az első mutagének, amelyeket Szaharov, Lobashev és alkalmazottai felfedeztek, alacsony hatékonysággal rendelkeztek, így nem tudtak érdeklődni a szakemberek számára.
A kémiai tényezőknek a mutációs folyamatban betöltött szerepének tanulmányozásában egy új szakaszt nyitottak meg I.A. Rapoport (1943,46,47) egyes vegyi anyagok erős mutagén hatását jelezte. Szintén széleskörű munkát szervezett a kémiai mutagének felhasználására a tenyésztés gyakorlatában.
Következtetés 4. szakasz
Az orosz tudósok a világon elsők között kezdték meg a mutációk mesterséges megszerzésére irányuló kísérleteket, ami lehetővé tette számukra, hogy következtetéseket vonjanak le a szervezetek variabilitását befolyásoló tényezőkről, az új szervezetek gyakorlati alkalmazásának lehetőségéről és a mutációk lehetséges következményeiről.
5 A GENE BRIBILITY PROBLÉMA
A XX. Század 30-as évek elejére. a gén elméletének alapját képezte. A hibridológiai elemzés első eredményei már felvetették az örökletes anyagok diszkrétségének problémáját. Úgy vélték, hogy a gén felelős az egyetlen tulajdonság kifejlesztéséért, és átadással, oszthatatlan egészként kerül átadásra. A mutációk és az átkelés felfedezése (a géneknek a kromoszómák közötti szakaszok közötti cseréje következtében bekövetkező kapcsolatának megszakítása) megerősítette a gének oszthatatlanságát. Az összes adat általánosítása eredményeként a gén definíciója a következő készítményt kapta: a gén az öröklődés elemi egysége, amelyet egy teljesen határozott funkció jellemez, amely az átkelés során egészében mutálódik. Más szóval, a gén a genetikai funkció, a mutáció és az átkelés egysége.
1928-ban az A.S. Serebrovsky a Biológiai Intézetben. KA Timiryazeva N.P. Dubinin elkezdte vizsgálni a röntgensugárzás hatását a Drosophilán, és felfedezett egy szokatlan mutációt. A kocka kialakulását egy repülés testén egy speciális gén szabályozza pikkely . Génmutáció pikkely Az amerikai genetikus, Payne (1920) által először felfedezett kísérletek többször is megjelentek a kísérletekben, és amikor megjelent, kilenc állomány kialakulását elnyomta. A Dubinin által feltárt mutáció elnyomta mindössze négy fajta fejlődését. További kísérletek után világossá vált, hogy a gén nem osztható genetikai struktúra, hanem a kromoszóma egy része, amelynek egyes részei egymástól függetlenül mutálhatnak. Ez a jelenség lépésenként allélomorfizmus.
Az osztályozott allélmorfok vizsgálatának egyik fő előnye a mutánsok számolásának kvantitatív módszere volt. Egy olyan rendszer kifejlesztése, amely lehetővé teszi az egyes mutációk eredményének számszerűsítését, Serebrovsky, Dubinin és más szerzők egyidejűleg felfedték az egyik mutáns gén hozzáadásának jelenségét. Ezt a jelenséget ezután újra felfedezték a mikroorganizmusokon, és megkapta a komplementáció nevét.
A gén mutációs törékenységét Serebrovszkij és laboratóriumának más alkalmazottai azonban már régóta nem tudták megerősíteni a gén fragmentálódását átkelés segítségével. A géntörés kimutatásához szükség volt egy hatalmas legyek ellenőrzésére. Ilyen kísérletet csak 1938-ban lehetett megszervezni, amikor Dubinin, N.N. Sokolov és G.G. Tinyakov képes megtörni a gént pikkely és ellenőrizze eredményét a Drosophila nyálmirigyének óriás kromoszómáiban.
Következtetés 5. szakasz
Bár a végső döntés arról, hogy a gént nemcsak mutánsan, hanem mechanikusan osztjuk-e meg, és hogy az orosz tudósok munkájában nem sikerült, jelentősen hozzájárultak a génmegosztottság bizonyításához, ami a mikroorganizmusok genetikai kutatásához, és ezáltal a genetika átmenetéhez vezetett. a kutatás molekuláris szintjén.
6 MOLEKULÁRIS GENETIKA
A gén fő funkciójának megismerése egy bizonyos polipeptidlánc szerkezetére vonatkozó információ tárolójaként felvetette azt a kérdést, hogy a genetika miként továbbítja az információt a genetikai struktúrákból (DNS) a morfológiai struktúrákba, vagyis a genetikai információ rögzítésének módjába és annak megvalósításába egy sejtben.
A Watson-Crick modell szerint a DNS genetikai információi hordoznak
bázisok szekvenciája. Így a DNS négy genetikai információ elemet tartalmaz. Ugyanakkor 20 esszenciális aminosavat találtak a fehérjékben. Szükség volt arra, hogy megtudjuk, hogyan lehet a DNS-ben egy négybetűs nyelvet lefordítani egy húsz betűs nyelvre Beksben. A mechanizmus kifejlesztésében döntő szerepet játszott G. Gamow. Azt javasolta, hogy három DNS-nukleotid kombinációját alkalmazzuk egyetlen aminosav kódolására (egy vegyület, amely cukorból (deoxoribóz), foszfátból és bázisból áll és alkotórész monomer DNS-t képez) nevezzük nukleotidnak. Ezt az elemi anyagot, amely egyetlen aminosavat kódol, kodonnak nevezzük.
Következtetési szakasz hat
Ebben a szakaszban az egyik orosz tudós fontos lépést tett a genetikai kód megoldása felé.
KÖVETKEZTETÉS
E munka anyagai alapján arra a következtetésre juthatunk, hogy az orosz tudósok hozzájárulása a genetika fejlődéséhez, mint tudomány, meglehetősen nagy, de alulértékelt. Ezt magyarázza a nagyrészt ismeretlen orosz tudományos folyóiratok, amelyek közzétették a külföldi tudósok munkáját.
Az orosz tudósok által végzett felfedezések jelentősége azonban vitathatatlan.
Meglehetősen nehéz leírni az orosz genetikusok által végzett munkát, mivel kísérleteik összetettek és változatosak. E munka keretében csak a tudomány további fejlődését tükröző kulcsfontosságú felfedezések tükröződnek.
BIBLIOGRAPHIC LIST
1. Új orosz enciklopédia: 12 t. / Redkol: A. D. Nekipelov, V. I. Danilov-Danilyan és mások - Moszkva: Enciklopédia, 2007. T. 3: Brunei - Vincha. - 480 s.
2. Biológia - A genetika fejlődésének története. - http://www.5ka.ru/10/24545/1.html
3. Wikipedia - Genetika. - http://en.wikipedia.org/wiki/Genetics
4. Wikipedia - Nikolay Ivanovich Vavilov. - http://ru.wikipedia.org/wiki/Vavilov,_Nikolai_Ivanovich
5. Nikolay Koltsov. - http://idbras.comcor.ru/personal/kolzov.htm
6. Vladimir Szaharov. - http://idbras.comcor.ru/personal/SakharovVVr.html
7. Nadson George Adamovich. - http://persona.rin.ru/view/f/0/23965/nadson-georgij-adamovich
8. A huszadik század nagy tudósai - Nikolai Vladimirovics Timofeev-Resovsky. - http://iomn.net/?p\u003d79
 A genetika történetének főbb időpontjai
A genetika történetének főbb időpontjai Egy barát vagy barátnő megjelenésének leírása
Egy barát vagy barátnő megjelenésének leírása Csillagok - a gyerekek magyarázata
Csillagok - a gyerekek magyarázata