هل يتم تحديد جنس الإناث دائمًا عن طريق مزيج من xx. بيولوجيا الأرض
علم الوراثة من الأرض
يتميز النوع الاجتماعي بمجموعة معقدة من السمات التي تحددها الجينات الموجودة في الكروموسومات. في خلايا الجسم البشري ، يتم الجمع بين الكروموسومات ومجموعات ثنائية الصبغيات. في الأنواع التي تحتوي على أفراد ثنائي الجنس ، لا يكون مركب الكروموسوم من الذكور والإناث متشابها ويختلف من زوج واحد من الكروموسومات (الكروموزومات الجنسية). كانت الصبغيات المتطابقة لهذا الزوج تسمى X (X) - كروموسوم ، غير مزاوج ، غائبة عن الجنس الآخر - Y (play) - كروموسوم. بقية التي لا توجد اختلافات هي autosomes (A).
تحتوي الخلايا الأنثوية على اثنين من الكروموسومات الجنسية المتطابقة ، تم تحديدها XX ، في الرجال التي يتم تمثيلها من قبل اثنين من الكروموسومات X و Y. وبالتالي ، فإن مجموعة الكروموسومات للرجل والمرأة تختلف فقط في كروموسوم واحد: مجموعة الكروموسوم للمرأة تحتوي على 44 autosomes + XX ، والرجال - 44 autosomes + XY.
خلال تقسيم ونضج الخلايا الجرثومية في البشر ، يتم تكوين أمشاج ذات عدد أحادي الكروموسومات: البيض ، كقاعدة عامة ، يحتوي على كروموسومات 22 + X. وهكذا ، في النساء ، يتم تكوين نوع واحد فقط من الأمشاج (أمشاج مع الكروموسوم X). في الذكور ، تحتوي الأمشاج على كروموسومات 22 + X أو 22 + Y ، ويتم تكوين نوعين من الأمشاج (مشيج مع كروموسوم X وشيجيت مع كروموسوم Y). إذا أدخلت الحيوانات المنوية مع الكروموسوم X ، أثناء الإخصاب ، خلية البويضة ، يتم تكوين جرثومة أنثى ، ومع كروموسوم Y يكون الذكر.
لذلك ، فإن تحديد جنس الشخص يعتمد على وجود الخلايا الجنسية الذكرية - الحيوانات المنوية ، أو تخصيب البويضة ، أو X- أو الكروموسومات Y.
هناك أربعة أنواع رئيسية من تحديد الجنس الكروموسومي:
1. الجنس الذكري هو متغاير. 50٪ من الأمشاج يحمل X- ، 50٪ -و- كروموسوم ، على سبيل المثال ، الإنسان ، الثدييات ، dipterans ، الخنافس ، الخلل (شريحة 4).
2. جنس الذكر هو heterogametic. 50٪ من الأمشاج هي X- ، 50٪ ليس لديهم كروموسوم جنسي ، على سبيل المثال ، جنادب ، كناغر (شريحة 7).
3. الجنس الأنثوي هو heterogametic. 50٪ من الأمشاج يحمل X- ، 50٪ g gamete –Y- كروموسوم ، على سبيل المثال ، الطيور ، الزواحف ، البرمائيات الذيل ، دودة القز (شريحة 7).
4. الجنس الأنثوي هو heterogametic. 50٪ من الأمشاج يحمل X- ، 50٪ ليس لديهم كروموسوم جنسي ، على سبيل المثال ، مول.
يسمى توارث الصفات التي يتم تحديد جيناتها في الكروموسومات الجنسية بالوراثة ، إلى جانب الكلمة.
26- النمط الجيني كنظام متكامل. تفاعل الجينات ، الفعل المتعدد للجينات.
النمط الجيني مثل نظام كلي
خصائص الجينات. على أساس أبحث مع أمثلة لميراث الصفات في الصلبان الأحادية والهجينة ، قد يحصل المرء على الانطباع بأن النمط الجيني للكائن يتكون من مجموع جينات منفصلة مستقلة التمثيل ، يحدد كل منها تطور سمة أو خاصية خاصة به. إن مثل هذه الفكرة عن ارتباط مباشر لا لبس فيه لجين ذي سمة غالباً لا تتوافق مع الواقع. في الواقع ، هناك عدد هائل من علامات وخصائص الكائنات الحية ، التي يحددها زوجان أو أكثر من الجينات ، والعكس صحيح ، فإن أحد الجينات يتحكم في كثير من الأحيان في العديد من العلامات. بالإضافة إلى ذلك ، يمكن تغيير تأثير الجين عن قرب الجينات الأخرى والظروف البيئية. وهكذا ، في الجين ، لا تعمل الجينات الفردية ، ولكن النمط الجيني كله كنظام متكامل مع اتصالات معقدة وتفاعلات بين مكوناته. هذا النظام ديناميكي: ظهور الأليلات أو الجينات الجديدة نتيجة للطفرات ، يؤدي تكوين كروموسومات جديدة وحتى جينومات جديدة إلى تغير ملحوظ في النمط الجيني بمرور الوقت.
قد تختلف طبيعة مظهر عمل الجينات في تكوين النمط الجيني كنظام في حالات مختلفة وتحت تأثير عوامل مختلفة. هذا يمكن رؤيته بسهولة إذا ما أخذنا في الاعتبار خصائص الجينات وخصائص مظاهرها في العلامات:
الجين منفصل في عمله ، بمعنى أنه معزول في نشاطه من جينات أخرى.
الجين محدد في مظهره ، أي أنه مسؤول عن خاصية محددة تمامًا أو خاصية الكائن الحي.
يمكن للجين أن يتصرف تدريجيًا ، أي زيادة درجة إظهار سمة مع زيادة في عدد الألائل السائدة (الجينات الجينية).
يمكن لجين واحد أن يؤثر على تطور السمات المختلفة - وهذا تأثير متعدد أو ثنائي المظاهر لجين.
يمكن للجينات المختلفة أن يكون لها نفس التأثير على تطور نفس الصفات (السمات الكمية في كثير من الأحيان) - فهي عبارة عن جينات متعددة أو مضادات متعددة.
يمكن للجين التفاعل مع جينات أخرى ، مما يؤدي إلى ظهور علامات جديدة. يتم مثل هذا التفاعل بشكل غير مباشر - من خلال منتجات ردود أفعالهم المركبة تحت سيطرتهم.
يمكن تعديل عمل الجين عن طريق تغيير موقعه على الكروموسوم (التأثير الموضعي) أو بتأثير العوامل البيئية المختلفة.
تفاعلات الجينات الأليلية. وتسمى هذه الظاهرة عندما تسمى العديد من الجينات (alleles) المسؤولة عن سمة واحدة التفاعل الجيني. إذا كانت هذه هي الأليلات من نفس الجين ، ثم تسمى هذه التفاعلات أليلية، وفي حالة الأليلات من جينات مختلفة - غير الأليلي.
تتجلى الأنواع الرئيسية التالية من التفاعلات الأليلية: الهيمنة ، والهيمنة غير المكتملة ، والإفراط في الدراسة ، والسودانية.
هيمنة نوع التفاعل بين اثنين من الأليلات من جين واحد ، عندما واحد منهم يلغي تماما مظهر الآخر. هذه الظاهرة ممكنة في الحالات التالية: 1) الأليل السائد في الحالة المتغايرة يفسح المجال لتخليق المنتجات الكافية لإظهار سمة من نفس النوعية كما هو الحال في حالة الزيجوت الزيجوتي السائد في الشكل الأم ؛ 2) الأليل المتنحي غير نشط تماما ، أو منتجات نشاطه لا تتفاعل مع منتجات نشاط الأليل السائد.
أمثلة على هذا التفاعل من الجينات الأليلية هي هيمنة الأرجواني اللون أزهار البازلاء على الأبيض ، شكل بذور ناعمة على التجاعيد ، الشعر الداكن على ضوء ، عيون بنية فوق اللون الأزرق في البشر ، إلخ.
هيمنة غير كاملة أو طابع وسيط من الميراث لوحظ في الحالة عندما يختلف النمط الظاهري للهجين (متغاير الزيجوت) عن النمط الظاهري لكل من متماثل الزيجوت الوالدي ، أي أن التعبير عن السمة هو وسيط ، مع انحراف أكثر أو أقل نحو واحد أو الوالد الآخر. آلية هذه الظاهرة هي أن الأليل المتنحي غير نشط ، ودرجة نشاط الأليل المهيمن غير كافية لتوفير المستوى المرغوب من مظاهر السمة السائدة.
مثال على الهيمنة غير الكاملة هي الميراث. اللون الزهور في النباتات من جمال الليل (الشكل 3.5). كما يتبين من الرسم البياني ، فإن النباتات المتماثلة الزيجوت إما حمراء (AA) إما أبيض (أأ) الزهور ، و heterozygous (أأ) - وردي. عند عبور النباتات مع الزهور الحمراء والنباتات مع الزهور البيضاء في F 1 ، جميع النباتات لديها الزهور الوردية ، وهذا هو ، هناك طابع وسيط من الميراث. عند عبور الهجينة مع الزهور الوردية في F 2 هناك مصادفة من الانقسام من النمط الظاهري والنمط الوراثي ، منذ الزيجوتية السائدة (AA) مختلفة من الزيجوت المتغايرة (أأ). لذلك ، في هذا المثال مع النباتات الجمال ليلا ، وتقسيم في F 2 لون الزهرة هو عادة ما يلي - 1 أحمر (أأ): 2 وردي (أأ): 1 أبيض (أأ).
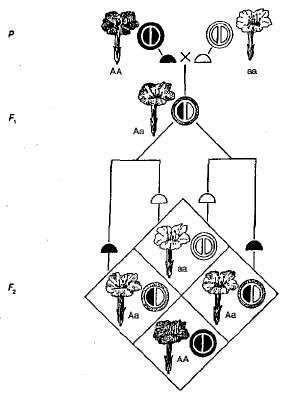
التين. 3.5. وراثة لون الزهرة في حالة الهيمنة غير المكتملة لجمال الليل.
أصبحت الهيمنة غير المكتملة واسعة الانتشار. ويلاحظ في ميراث الشعر المجعد في البشر ، ولون الماشية ، ولون الريش في الدجاج ، والعديد من الشخصيات المورفولوجية والفسيولوجية الأخرى في النباتات والحيوانات والبشر.
superdominance - مظاهر أقوى للسمات في شخص متغاير الزيجوت (أأ)، من أي زيجوت متماثل (AA و أأ). من المفترض أن هذه الظاهرة هي السبب في وجود تجلط الدم (انظر الفقرة 3.7).
Kodaminirvanie- مشاركة كلا الأليلين في تحديد السمات في فرد متماثل. من الأمثلة المدهشة والمدروسة جيداً على التزاوج المشترك هو وراثة مجموعة الدم الرابعة لدى البشر (المجموعة AB).
تحتوي خلايا الدم الحمراء للأشخاص في هذه المجموعة على نوعين من المستضدات: المستضد A (يحدده الجينوم / \\ متوفر في واحد من الكروموسومات) والمستضد ال (يحددها الجينوم / أ ، مترجمة على كروموسوم آخر متجانس). فقط في هذه الحالة الأليلات - 1 A (في الحالة متماثلة الزيجوت يتحكم في مجموعة الدم II ، المجموعة A) و أنا B (في الدولة متماثل الزيجوت III مجموعة الدم ، المجموعة ب). الأليلات 1 A و أنا B العمل في متغاير الزيجوت كما لو كان بشكل مستقل عن بعضها البعض.
الوراثة سبيل المثال مجموعة يوضح الدم ومظهره allellism متعددة: الجينات / يمكن تمثيلها بثلاثة ألليلات مختلفة ، وهناك جينات لديها عشرات من الأليلات. يتم تسمية جميع الأليلات من نفس الجين سلسلة من الأليلات المتعددة ، يمكن لكل كائن ثنائي الصبغيات أن يحتوي على أيلين (وفقط). بين هذه الأليلات ، كل المتغيرات المذكورة من التفاعلات الأليلية ممكنة.
ظاهرة التعددية المتعددة شائعة في الطبيعة. هناك سلسلة واسعة من الأليلات المتعددة التي تحدد نوع التوافق أثناء التخصيب في الفطريات ، التلقيح في نباتات البذور ، تحديد لون شعر الحيوان ، إلخ.
تفاعلات الجينات غير المتوازية تم وصف تفاعلات جين اللاأليل في العديد من النباتات والحيوانات. أنها تؤدي إلى ظهور في نسل digerozygotes من انشقاق غير عادي وفقا لظاهرة ظاهرية: 9: 3: 4؛ 9: 6: 1 ؛ 13: 3 12: 3: 1 ؛ 15: 1 ، أي تعديلات الصيغة العامة منديلي 9: 3: 3: 1. هناك حالات من التفاعل بين جينات اثنين ، ثلاثة وأكثر غير متوازية. ومن بين هذه الأنواع الرئيسية التالية: التكامل ، epistasis والبوليمرات.
مكملة، أو إضافية، ويسمى هذا التفاعل الجينات السائدة غير المتوازية ، ونتيجة لذلك تظهر سمة غير موجودة في كلا الوالدين. على سبيل المثال ، عندما يتم عبور نوعين من البازلاء الحلوة مع زهور بيضاء ، تظهر ذرية مع زهور أرجوانية. إذا قمت بتعيين النمط الجيني لمجموعة متنوعة واحدة AABB، والآخر aABB، ال
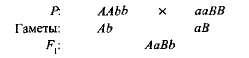
الجيل الأول الهجين مع اثنين من الجينات السائدة (A و في) حصل على أساس كيميائي حيوي لإنتاج الصباغ الأرجواني anthocyanin ، في حين أن واحدا تلو الآخر الجين و، لم يقدم الجين B تركيب هذا الصباغ. إن توليف الأنثوسيانين هو سلسلة معقدة من التفاعلات البيوكيميائية المتسلسلة التي تسيطر عليها العديد من الجينات غير المتوازية ، وفقط إذا كان هناك جينات جهازيتين على الأقل. (A-B) يتطور اللون الأرجواني. في حالات أخرى (aaV- و A-ب) زهور النبات بيضاء (العلامة "-" في صيغة التركيب الوراثي تعني أن هذا المكان يمكن أن يشغله كل من الأليل المسيطر والأليل المتنحي).
عندما النباتات البازلاء الذاتية التلقيح من F 1 في F 2 وقد لوحظ تقسيم إلى أشكال الأرجواني والأزهار البيضاء في نسبة قريبة إلى 9: 7. تم العثور على الزهور الأرجواني في 9/1 6 نباتات ، بيضاء - في 7/16. تظهر شعرية بينيت بوضوح سبب هذه الظاهرة (الشكل 3.6).
قشوة - هذا نوع من التفاعل الجيني تقوم فيه جينات أحد الجينات بقمع مظهر زوج جينات أليل من جينة أخرى. الجينات تمنع عمل الجينات الأخرى تسمى مثبطات إبيستاتيك أو المكثفات. يسمى الجين المكبوت صاغر.
حسب التغير في عدد ونمط الصفات الظاهرية والطبقات المضغوطة أثناء الانقسام الهجين في F 2 النظر في عدة أنواع من التفاعلات الاستعمارية: المهيمنة المهيمنة (A\u003e B أو B\u003e A) مع التقسيم 12: 3: 1 ؛ epistasis المتنحية (أ) ب أو ب \u003e أ) ، والتي يتم التعبير عنها في التقسيم 9: 3: 4 ، إلخ.
polymerism يتجلى في حقيقة أن يتم تشكيل علامة واحدة تحت تأثير العديد من الجينات مع نفس التعبير المظهري. تسمى هذه الجينات البوليمر. في هذه الحالة ، تم اعتماد مبدأ العمل الجيني للجين على تطوير السمات. على سبيل المثال ، عند عبور نباتات كيس الراعي مع ثمار مثلثية وبيضاوية (Pods) في F 1 ، يتم تكوين نباتات ذات ثمار مثلثة الشكل. مع التلقيح الذاتي في F 2 وينقسم إلى النباتات مع القرون الثلاثي والبيضاوي في نسبة 15: 1 لوحظ. ويفسر ذلك حقيقة أن هناك جينين يعملان بشكل فريد. في هذه الحالات ، يتم دلالة مماثلة. A 1 و A 2 .
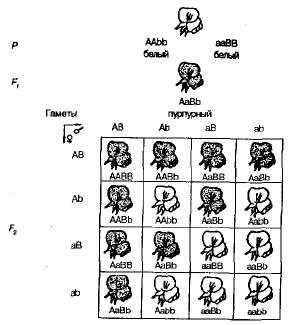
التين. 3.6 . وراثة لون الزهرة في البازلاء الحلوة
ثم كل الأنماط الجينية (A 1 ,-A 2 ,-, A 1 الصورة 2 و 2 و 1 ل 1 A 2 -) سيكون له نفس النمط الظاهري - القرون الثلاثي ، والنباتات فقط و 1 و 1 و 2 ل 2 سوف تكون مختلفة - لتشكيل قرون البيضاوي. هذا هو الحال بوليمرات غير تراكمية.
يمكن للجينات البوليمرية التصرف حسب النوع البوليمرات التراكمية. الجينات الأكثر تشابهاً في النمط الجيني للكائن الحي ، كلما كان مظهر هذه السمة أقوى ، أي بزيادة جرعة الجين. (A 1 A 2 A 3 إلخ) يتم تلخيص الإجراء الخاص به أو تجميعه. على سبيل المثال ، تتناسب شدة لون سويداء حبيبات القمح مع عدد الألائل السائدة من جينات مختلفة في العبور الثلاثي الهجين. وكانت معظم الحبوب الملونة A 1 A 1 A 2 A 2 A 3 أ 3 و الحبوب 1 و 1 و 2 ل 2 و 3 و 3 لم يكن لديك الصباغ.
وفقا لنوع البوليمر التراكمي ، هناك العديد من العلامات الموروثة: اللبن ، إنتاج البيض ، الوزن وعلامات أخرى لحيوانات المزرعة. العديد من البارامترات الهامة للقوة البدنية للشخص ، والقدرات الصحية والعقلية ؛ طول أذن الحبوب. محتوى السكر في جذور بنجر السكر أو الدهون في بذور عباد الشمس وهلم جرا
وبالتالي ، فإن العديد من الملاحظات تشير إلى أن مظهر معظم العلامات هو نتيجة لتأثير مركب من الجينات المتفاعلة والظروف البيئية على تكوين كل سمة محددة.
التفاعل الجيني
العلاقة بين الجينات والصفات معقدة للغاية. في الكائن الحي ، لا يحدد الجين واحدًا سمة واحدة فقط ، وبالعكس ، لا يتم تحديد سمة إلا بواسطة جين واحد. في كثير من الأحيان ، يمكن أن تساهم جينة واحدة في إظهار عدة علامات في وقت واحد ، والعكس بالعكس. لا يمكن اعتبار النمط الجيني للكائن الحي كمجموع بسيط من الجينات المستقلة ، يعمل كل منها على اتصال بالآخرين. المظاهر المظهريّة لسمات ما هي نتيجة تفاعل العديد من الجينات.
العمل الجيني المتعدد (pleiotropy) - عمليات تأثير جين واحد على تشكيل العديد من الشخصيات.
على سبيل المثال ، في البشر ، الجين الذي يحدد لون الشعر الأحمر يسبب بشرة ونمش أخف.
في بعض الأحيان ، تؤثر الجينات التي تحدد الخصائص المورفولوجية على الوظائف الفسيولوجية ، مما يقلل من الحيوية والخصوبة ، أو تكون مميتة. وهكذا ، فإن الجين الذي يسبب لونًا أزرقًا في المنك يقلل من خصوبته. تم تفصيل الجين السائد للتلوين الرمادي في خراف العرقول في الحالة المتماثلة الزيجوت ، لأن هذه الحملان تعاني من نقص في المعدة وتموت عندما تتحول إلى عشب.
تفاعل الجينات التكميلية. يمكن لعدة جينات التأثير على تطوير سمة واحدة. يسمى تفاعل العديد من الجينات غير المتوازية ، مما يؤدي إلى تطوير سمة واحدة ، باسم تكاملية. على سبيل المثال ، تحتوي الدجاج على أربعة أشكال من قمة ، ويرتبط مظهر أي منها بتفاعل اثنين من أزواج الجينات غير المتوازية. يعود الفضل في هذه الوردة الوردية إلى عمل الجين السائد لأليل واحد ، وهو قمة على شكل البازلاء - الجين السائد للأليل الآخر. في الهجينة ، في وجود اثنين من الجينات غير المتوازية المهيمنة ، يتم تشكيل قمة تشبه الجوز ، وفي غياب جميع الجينات السائدة ، أي في متماثل الزيجوتية المتنحية لاثنين من الجينات غير المتوازية ، يتم تشكيل مشط بسيط.
نتيجة تفاعل الجينات هي تلوين الصوف في الكلاب ، والفئران ، والخيول ، وشكل اليقطين ، ولون أزهار البازلاء الحلوة.
البوليمر هو تفاعل بين جينات غير متوازية عندما تعتمد درجة تطور سمة ما على العدد الكلي للجينات السائدة. وفقا لهذا المبدأ ، يتم توارث لون الشوفان وحبوب القمح ولون الجلد للشخص. على سبيل المثال ، يوجد في السود 4 جينات مسيطرة في زوجين من الجينات غير المتوازية ، ولا يوجد في البشر ذوي البشرة البيضاء ، كل الجينات تنحية. مزيج من كميات مختلفة من المهيمنة و جينات متنحية يؤدي إلى تشكيل mulattos مع كثافة مختلفة من لون البشرة: من الظلام إلى الضوء.
هناك مجموعتان رئيسيتان من التفاعل الجيني: التفاعل بين الجينات الأليلية والتفاعل بين الجينات غير الأليلية. ومع ذلك ، ينبغي أن يكون مفهوما أن هذا ليس التفاعل المادي للجينات نفسها ، ولكن التفاعل بين المنتجات الأولية والثانوية التي ستحدد سمة واحدة أو أخرى. في السيتوبلازم ، يحدث التفاعل بين البروتينات - الإنزيمات التي يتم تحديد تركيبها عن طريق الجينات ، أو بين المواد التي تتشكل تحت تأثير هذه الإنزيمات.
الأنواع التالية من التفاعل ممكنة:
1) لتكوين سمة محددة ، يكون تفاعل اثنين من الإنزيمات ضروريًا ، يتم تحديد تركيبه عن طريق جينات غير أليل ؛
2) إن الإنزيم الذي تم تصنيعه بمشاركة جينة واحدة يكبت أو يثبط نشاط الإنزيم الذي يتكون من جين آخر غير متوازي ؛
3) إنزيمين ، يتم التحكم في تكوينهما بواسطة جينيْن غير أليلين يؤثران على سمة واحدة أو عملية واحدة بحيث يؤدي عملهما المشترك إلى ظهور وتعزيز مظهر السمات.
تفاعل الجينات الأليلية
تسمى الجينات التي تشغل مواقع متماثلة (متماثلة) في الكروموسومات المتماثلة باللاتينية. كل كائن حي لديه جينات ألين.
ومثل هذه الأشكال من التفاعل بين الجينات الأليلية معروفة: الهيمنة الكاملة ، والهيمنة غير المكتملة ، والاقتران المفرط ، والسيطرة المفرطة.
الشكل الرئيسي للتفاعل هو الهيمنة الكاملة ، التي وصفها G. Mendel لأول مرة. يكمن جوهرها في حقيقة أنه في كائن حي متخالف يتجلى مظهر أحد الأليلات على مظهر الآخر. مع الهيمنة الكاملة على التقسيم عن طريق التركيب الوراثي 1: 2: 1 لا يتطابق مع التقسيم عن طريق النمط الظاهري - 3: 1. في الممارسة الطبية ، من ألفي من الأمراض الوراثية أحادية الجين في النصف تقريبا ، تحدث مظاهر مسيطرة على الجينات المرضية على وضعها الطبيعي. في متغاير الزيجوت ، يتجلى في الأليل المرضية في معظم الحالات من علامات المرض (النمط الظاهري السائد).
الهيمنة غير المكتملة هي شكل من أشكال التفاعل الذي لا يقوم فيه الجين السائد (A) في كائن متجانس (Aa) بقمع الجين المتنحي (أ) بالكامل ، ونتيجة لذلك يظهر وسيط بين العلامات الأبوية. هنا يتقسم التقسيم في النمط الوراثي والنمط الظاهري وهو 1: 2: 1
عندما تتحول العوامل الوراثية في الكائنات المتغايرة ، فإن كل جين من الجينات الأليلية يتسبب في تكوين منتج يعتمد عليه ، أي أن نتاج كلا الأليلين يكون. أحد الأمثلة الكلاسيكية لمثل هذا المظهر هو نظام مجموعة الدم ، ولا سيما نظام ABO ، عندما تحمل كريات الدم الحمراء البشرية مستضدات على السطح تتحكم فيها كلتا الأليلات. يسمى هذا الشكل من مظاهر التزاوج.
فرط - عندما يكون الجين السائد أكثر وضوحا في حالة الزيجوت المتغاير عنه في الحالة المتماثلة الزيجوت. وهكذا ، في الدروسوفيلا ، مع النمط الجيني AA ، العمر المتوقع الطبيعي ؛ أأ - حياة trivatist الممتدة. أأ - قاتلة.
المزيفة متعددة
كل كائن حي لديه جينات ألين فقط. ومع ذلك ، في كثير من الأحيان ، يمكن أن يكون عدد الأليلات أكثر بطبيعتين ، إذا كان بعض المواقع يمكن أن يكون في حالات مختلفة. في مثل هذه الحالات ، يقال متعددة الأليلات أو متعددة الأشكال.
يتم تحديد الأليلات المتعددة بحرف واحد مع مؤشرات مختلفة ، على سبيل المثال: A ، A1 ، A3 ... توجد جينات Allelic في نفس المناطق من الكروموزومات المتجانسة. بما أن هناك دائما كروموسومين متشابهين في النمط النووي ، حتى مع الألائل المتعددة ، يمكن لكل كائن حي أن يكون له فقط أليلين متماثلين أو مختلفين في وقت واحد. يدخل واحد منهم فقط الخلية التناسلية (جنبا إلى جنب مع الاختلاف في الكروموسومات المتماثلة). لللاتيلات المتعددة ، التأثير المميز لجميع الأليلات على نفس الصفة. الفرق بينهما لا يكمن إلا في درجة تطور الصفة.
الميزة الثانية هي أن الخلايا أو الخلايا الجسدية من الكائنات ثنائية الصبغيات تحتوي على ألحانين مختلفين كحد أقصى ، لأنها تقع في موضع الكروموسوم نفسه.
ميزة أخرى متأصلة في الأليلات المتعددة. وبحكم طبيعة الهيمنة ، يتم وضع الأحرف المتجاورة في صف متسلسل: فغالبًا ما تهيمن السمة الطبيعية غير المتغيرة على السمات الأخرى ، أما الجين الثاني من السلسلة فهو مقهور بالنسبة للأول ، ولكنه يهيمن على ما يلي ، وما إلى ذلك. أحد الأمثلة على مظهر الألائل المتعددة في الشخص هو مجموعات الدم في نظام ABO.
تتميز المراتب المتعددة بأهمية بيولوجية وعملية مهمة ، لأنها تزيد من التقلبات التوافقية ، وخاصة النمط الجيني.
تفاعل الجينات غير allel
هناك العديد من الحالات التي يتم فيها تحديد سمة أو خصائص من قبل اثنين أو أكثر من الجينات التي لا يمكن فصلها تتفاعل مع بعضها البعض. على الرغم من أن التفاعل هنا مشروط ، لأنه ليس الجينات التي تتفاعل ، ولكن المنتجات التي يسيطرون عليها. في هذه الحالة ، هناك انحراف عن أنماط تقسيم Mendelian.
هناك أربعة أنواع رئيسية من التفاعل الجيني: التكامل ، و epistasis ، والبوليمرات ، وتعديل العمل (pleiotropy).
والتكامل هو نوع من التفاعل بين جينات غير متوازية ، عندما يكمل أحد الجينات المهيمنة عمل جين مهيمن آخر غير متوازي ، ويحدِّد معاً سمة جديدة غير موجودة في الوالدين. علاوة على ذلك ، فإن السمة المناظرة تتطور فقط في وجود كل من الجينات غير الأليلية. على سبيل المثال ، الكبريت هو لون الصوف في الفئران التي تسيطر عليها جيناتين (A و B). يحدد جين A تركيب توليف الصبغ ، ومع ذلك ، فإن كلا من مضادات الزيجوت (AA) والأجنة المتغايرة (Aa) هي ألبينو. ويقدم جين آخر (ب) مجموعات صبغية أساسًا في القاعدة وعلى أطراف الشعر. تهجين الديهيراتوزيوز (AaBb x AaBb) يؤدي إلى تقسيم الهجينة في نسبة 9: 3: 4. يمكن أن تكون النسب العددية للتفاعلات التكميلية في 9: 7 ؛ 9: 6: 1 (تعديل تقسيم Mendeleev).
مثال على التفاعل التكميلي للجينات في البشر يمكن أن يكون توليف بروتين وقائي ، مضاد للفيروسات. يرتبط تكوينه في الجسم بالتفاعل التكميلي لجينين غير أليلين يوجدان في كروموسومات مختلفة.
Epistasis هو تفاعل من جينات غير متوازية يقوم فيها أحد الجينات بقمع عمل جين آخر غير متوازي. يمكن لكل من الجينات السائدة والمُتَوَسِّرة أن تسبب الاكتئاب (A\u003e B ، A\u003e B ، B\u003e A ، B\u003e A) ، واعتماداً على هذا ، فإن المهارة هي المهيمنة والمنحية. يسمى الجين الساحق المانع أو القامع. عموما لا تحدد الجينات المانع تطوير سمة معينة ، ولكن فقط تمنع عمل جين آخر.
يسمى الجين الذي يتم قمع تأثيره بـ hypostatic. مع تفاعل الجينات الإستشاري ، يكون انقسام النمط الظاهري في F2 هو 13: 3 ؛ 12: 3: 1 أو 9: 3: 4 ، إلخ. لون ثمار اليقطين ، يتم تحديد لون الخيول بهذا النوع من التفاعل.
بول - وهي مجموعة من العلامات المورفولوجية والفيزيولوجية والبيوكيميائية والسلوكية وغيرها من علامات الجسم ، مما يسبب التكاثر.
وتنقسم العلامات التي يختلف فيها الأفراد من الجنسين المختلفين إلى: الابتدائي والثانوي. الأعضاء الأولية هي الأعضاء التي تضمن تكوين الأمشاج والإخصاب (الغدد التناسلية ، السبيل التناسلي ، الأعضاء). هذه هي الأعضاء التناسلية الخارجية والداخلية ، والتي وضعت في مرحلة التطور الجنيني. ثانوية - لا تشارك في التكاثر الجنسي. تطوير تحت تأثير الهرمونات الجنسية وتظهر في سن البلوغ (في البشر في 12-15 سنة). هذه هي ملامح تطوير الجهاز العضلي الهيكلي ، الأنسجة الدهنية تحت الجلد ، والشعر ، والجرس الصوتي ، والسمات السلوكية. في الطيور ، الغناء ، الريش ، إلخ.
يمكن تقسيم علامات الأفراد المرتبطين بالجنس إلى 3 فئات:
1) محدودة على الأرض
2) الكلمة التي تسيطر عليها
3) إلى جانب الكلمة.
ويرجع تطور الحالة الأولى إلى الجينات الموجودة في الجسيمات الذاتية لكلا الجنسين ، ولكنها تظهر فقط في جنس واحد. وهكذا ، فإن جينات وضع البيض موجودة في الدجاج والديكه ، ولكن تظهر فقط في الدجاج. وبالمثل ، تتصرف جينات الدهون والحليب في الماشية. ولوحظت هذه الظاهرة فيما يتعلق بالتعرض لهرمونات الجنس المقابلة.
مثال على الثاني هو مظهر من القرنية في الأبقار الذكور ، komolost - في الإناث. في البشر: الصلع في الذكور ، والنقرس - 80 ٪ في الذكور و 12 ٪ في الإناث.
تسمى العلامات ، التي يتحكم تطويرها بجينات الكروموسومات الجنسية ، بالجنس. هناك حوالي 200 علامة من هذا النوع ، مع وجود كروموسوم X هناك عمى لون مرتبط ، الهيموفيليا. Y-chromosome - فرط الشعر ، السماك.
أنواع تحديد الدور.
1. بروغاموس - قبل الإخصاب. لا تلعب نسبة الكروموسومات الجنسية في هذه الحالة دورًا البويضات ثنائية الصبغة. (بعض الديدان ، الروتيفر - تطور الإناث من البويضات الكبيرة ، الذكور من البويضات الصغيرة).
2. سيناميك - التحديد الجيني للجنس أثناء الإخصاب ، والذي يعتمد على طبيعة توليفة الكروموسومات الجنسية أو نسبة الكروموسومات الجنسية والجسيمات.
3. Epigamic - تحت تأثير البيئة الخارجية (دودة bonelia).
تعريف الكروموسومات من الطابق
┌─────────────────────────┬───────────────────┬──────────────────┐
│ أنواع الكروموسومات │ التراكيب الوراثية │ أنواع شبيه │
│ تحديد الجنس ├─────────┬─────────┼────────┬─────────┤
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
he heterogame │ │
│ حشرات اللقلق │ │ │ │ │
b (البق Protenor ، الخنافس ، │ │ │ │ │
│pauki، grasshoppers) │ ХО │ ХХ │ Х، О │ Х │
│ Drosophilus │ XY │ XX │ X، Y X │
│ الفقاريات │ │ │ │ │
│ (الثدييات ، والرجل) │ XY │ XX │ X ، Y │ X │
│ │ │ │ │ │
he أنثى متغايرة │ │
│ طيور. السمك ، الفراشات ، │ │ │ │ │
│ коп │ │ │ │
│ المائية. │ XX │ XY │ X │ X، Y │
andالسعر وغيرها من عاجزة │ │ │ │ │
Night │ XX │ XO │ X │ X، O
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
نوع التعريف syngamic هو تعريف الكروموسومات الجينات ذات السيطرة الجينية. مسؤول عن جنس الصبغي يسمى الجنس. يحمل المشيج الذكري العادي إما كروموسوم X أو Y ، وكل البيض - الكروموسوم X. في حالة تباين الكروموسوم الطبيعي أثناء الانقسام الاختزالي ، يتم تكوين البيض الطبيعي والحيوانات المنوية مع المجموعة المعتادة من الكروموسومات X و Y. يتم تحديد جنس الزيجوت بواسطة نسبة الأمشاج XX و XY (homogametic و heterogametic).
نظرية كروموسوم Correns للجنس (1907) هي أن الجنس يتم تحديده عن طريق الجمع بين الكروموسومات الجنسية أثناء الإخصاب. تتميز الأنواع التالية من تحديد جنس الكروموسومات: XY، XO، ZW، ZO.
في حالة انتهاك مسار الانقسام الفتيلي أو الانقسام الاختزالي ، يمكن تشكيل جينوروغوربس. قد يختلف محتوى الصبغيات الجنسية في خلايا مختلفة من هؤلاء الأفراد (الفسيفساء). في ذبابة ذبابة الفاكهة (Drosophila fly): XX و XO ، في الرجل XX و XY ، فيما يتعلق بآلام الجسم المختلفة التي يمكن أن تحمل علامات متشابهة للجنس. قد تكون هناك حالات أخرى من الفسيفساء: XX / XXX ، XY / XXX ؛ XO / XXY وغيرها
إذا لم تتفكك كروموسومات الجنس ، فقد يكون هناك 12 تركيبة ممكنة من الكروموزومات الجنسية في اللقاحات البشرية ، التي هي سبب الانحرافات الصبغية في البشر.
│ X │ XX │ O
─────┼────────┼─────────┼────────
X │ XX │ XXX │ XO
Y │ XY │ XXY │ YO
XY │ XXY │ XXXY │ XYO
O │ XO │ │
في حالة عدم وجود الصبغيات الجنسية أثناء الانقسام الاختزالي ، تتشكل الأمشاج XX و O في الإناث. و XY و O في الذكور. بمشاركة من الإخصاب ، تتشكل الجراثيم بمزيج غير عادي من الكروموزومات الجنسية. في البشر ، تحدث هذه الحالات الشاذة 1 في 600-700 حديثي الولادة. يموت Zygote يو في مرحلة مبكرة. الأفراد XXX ، XXY ، XO قابلة للحياة ويعتمد جنسهم على وجود أو عدم وجود الكروموسوم "Y" ، الذي ، مع أي عدد من الكروموسومات X ، يتحكم في تكوين علامات للجنس الذكر ، والتنمية ، ويحفز على تكوين الخصيتين. تسبب فائض من الكروموسومات X تشوهات دستورية وعيوب في الذكاء. ولكن في الطبيعة هناك أفراد يكون الكروموسوم "Y" خاملًا وراثيًا وليس له تأثير خاص على تحديد الجنس.
تم العثور على عينات ذبابة الفاكهة من نوع CW ، والتي كانت من الذكور ولكن العقم (1916 ، جيم الجسور) ، والأفراد XXY هم الإناث الخصبة العادية.
نظرية التوازن في الجنس (الجسور ، 1922). درس نسبة الأعضاء التناسلية و autosomes
في الإناث العادية مع مجموعة من الكروموسومات 2n ، تكون نسبة autosomes و X كروموسومات 1: 2n = 2A + 2X (2X: 2A = 1 - أنثى عادية) ، 1 ، 5 - فوق الإناث: 2A + 3X (3X: 2A = 1، 5 - عقيم). في الذكور ، تكون النسبة 0 ، 5 2n = 2A + XY (X: 2A = 0، 5). مع انخفاض فرده تبقى الذكور 3A + XY (X: 3A = 0 ، 33 - جرداء) - supermen. تتطابق قيمة المعامل بين 1 و 0 ، 5 مع النمط الظاهري للأفراد الوسيطة حسب الجنس _intersex: 3A + 2X (2X: 3A = 0 ، 66 - علامات كلا الجنسين عقيم).
وبالتالي ، فإن جوهر نظرية التوازن هو أن ليس فقط الكروموسومات الجنسية ولكن أيضا الجسيمات الصناعية المشاركة في تحديد الجنس. مجموعة أحادية الصبغيات أحادية الصيغة الصبغية تحكي خصائص الذكور الفردية. في هذه الحالة ، يتم تحديد الكلمة من خلال نسبة عدد (التوازن) من autosomes والصبغيات الجنسية.
تم العثور على تحديد الجنس من قبل ploidy أيضا في النحل. الإناث هي مضاعفة والذكر هم فرداني ، لأن تطوير المسببات العرقية من البيض غير المخصبة.
فروق الأرضيات في التضاريس
لا تختلف بدايات الغدد التناسلية في الأجنة المبكرة (حتى الأسبوع الخامس أو السادس) في مختلف الجنسين وتتكون من الطبقة الخارجية - القشرة الدماغية والطبقة الداخلية - النخاع ، ولا تحتوي على خلايا خط جرثومي. توجد الخلايا الجرثومية الأساسية لدى البشر في الأسبوع الثالث من التطور الجنيني في الأديم الظاهر من الكيس المحي. ثم ، تحت تأثير الإشارات chemotactic ، يهاجرون إلى الغدد التناسلية. هذه الهجرة مستقلة عن الجنس. يمكن أن تتطور أساسيات الغدد التناسلية في المبيضين أو الخصيتين. لوحظ التمايز التناسلية في الأسبوع الثامن: في اليوم 36 ، تبدأ الخصية بإفراز الأندروجين (التستوستيرون) ، الذي يحدد تطور الجنس الذكري.
في الكائنات الحية التي لها غدد جنسية ، تمارس السيطرة الجينية لتشكيل الخصائص الجنسية.
تتم برمجة التمايز بين الجنسين من خلال مجموعة وراثية من الكروموسومات الجنسية التي تشكلت خلال ملتقى الأمشاج الأنثوية والذكورية. يتم تحديد الجنس الوراثي للجنين بواسطة الكروموسومات XX أو XY.
يتم تحديد اتجاه تطور الجندر من خلال وجود كروموسوم Y. عادة ، تحتوي الكروموسومات X على جين معقم (Tfm ، جينات تأنيث الخصية) ، والذي يمنع تطور نوع الذكر. يحدد الأليل الطبيعي للجين تركيب مستقبل البروتين في الأندروجينات ، ويتم توليفه في كلا الجنسين. يعتمد تطور النمط الظاهري للذكور على جين الكروموسوم Y ، المستضد HY (الموصوف في الفأرة عام 1955 ؛ جين HA). يفرز من قبل الخلايا الجرثومية الذكور الابتدائية. Y 5NA 0 مسؤولة عن إنتاج هرمون التستوستيرون (هرمون الذكورة). حالما تصل هذه الخلايا إلى بدايات المناسل ، يبدأ تمييز الخصيتين. توجد مستقبلات HY على سطح كلا النوعين من خلايا الغدد التناسلية (الانحراف عن تطور الجنس في الأبقار). كان يعتقد أن النمط الظاهري الذكري يحدده كروموسوم الذكور بأكمله. ولكن في عام 1990 ، تم اكتشاف الجين (الجنس المنطقة Y) ، المترجمة في النمط النووي للكروموسوم Y. في غيابه ، يعطي النمط الوراثي XY النمط الظاهري الأنثوي.
تطوير الجندي
الغدد التناسلية الجرثومية المخنثين
│ ├┼┼┼┤ │ talnaya
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
إذا كان النمط الوراثي هو XX │ │ إذا كان النمط الوراثي هو XY
(7-8 أسابيع) ┴──┴──┐ ┌───┴───┐ (الأسبوع السادس)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
c └┴┘ │ │└┴┴┴┴┴┘│ الطبقة النخاعية القشرية
طبقة └─────┘ └───────┘
(يحدد الجين الجنس الأنثوي) (يحدد الجينات جنس الذكور)
في البشر ، الهرمونات الأنثوية المحددة التي تنظم وظائف
تصبح هذه العلامات ملحوظة ؛ فهي نتاج خلالي
لنا في نهاية 7-8 أسابيع. على نسيج 20 من خلية المبيض أو المنوية
الخلايا الجرثومية الأولية للخلية (خلايا Leydig). انها تشكل
الجنس الهرموني (التستوستيرون ،
الغدد التناسلية ومتباينة لاستراديول). 10-12 أسابيع لـ-
من oogonyev بنهاية الشهر الثاني. الكائنات الحية الداخلية
بحلول نهاية الشهر الثالث في العمق ، في الأسبوع الثاني عشر ، مع بعض
تم الكشف عن أجنة الغدد التناسلية على مستوى الأندروجينات في الجنين
البويضات (بروفس MI). يبدأ التفضيل التفاضلي (
الخراجات المبيض قبل سن 7 أشهر (صعود المتوسطة) ويكمل
وسوف تذوب أيضا بحلول الشهر التاسع في المبيض بحلول الأسبوع ال 20.
هناك 200-400000 البويضات الثاني. في سن البلوغ ، ومستوى
يزيد rogen ويؤثر
هيكل عظمي ، وكذلك
الاندروجين على التوالي (بواسطة
نوع الإناث والذكور).
يحدد الغدد التناسلية تطوير الخصائص الجنسية الأولية والثانوي. تفرز الغدد الجنسية الهرمونات (الاستراديول والأندروجينات) ، والتي ، جنباً إلى جنب مع هرمونات الغدد الصماء ، تتحكم في مسارات التمايز بين الجنسين. يتم التحكم في مستوى الهرمونات بدورها عن طريق الجينات.
وبالتالي ، فإن عملية التمايز الجنسي تشمل:
1) الرقابة الجينية ؛
2) الوظائف التنظيمية للهرمونات.
هناك نظرية لعمل الهرمونات كعوامل تنظيمية على الجينات. يتصرفون فقط على خلايا مستهدفة محددة. يتم إنتاج بروتين خاص في الخلية - وهو مستقبل يرتبط بالهرمون ويتغير في الوقت نفسه ، ثم يكتسب الخصائص لتحفيز عمل جينة واحدة أو عدة جينات في الكروموسومات. عندما يعمل الجلايكورتيكويدات على خلايا الرحم ، يتغير تخليق الحمض النووي الريبي والبروتينات فيها (انظر الرسم التخطيطي).
يتم التحكم في تكوين مستقبلات البروتين والهرمونات عن طريق الجينات. في حالة حدوث انتهاك للسيطرة ، من الممكن حدوث حالات شاذة ، مثال على ذلك هو متلازمة موريس. التأنيث الخصى (متلازمة موريس) (موريس ، 1953): مستقبلات التستوستيرون غائبة في الأفراد المصابين بهذا المرض. يتم إفراز الأندروجينات بكميات طبيعية. في متلازمة موريس ، تكون مرحلة التطور الجنيني هي أساس الخصيتين التي تبدأ في إنتاج هرمونات الذكورة الجنسية. ومع ذلك ، لا تشكل هذه الأجنة بروتينًا مستقبليًا (طفرة جينية متنحية) ، والذي يوفر حساسية خلايا الأعضاء النامية إلى هرمون الذكورة. بموجب هذا ، يتوقف نمو الذكور من النوع ويظهر النمط الظاهري الأنثوي. في حالات استثنائية ، من الممكن تصحيح هذه العيوب عن طريق إدخال هرمونات مناسبة.
وبالتالي ، فإن الإزدواجية الجينية الأصلية هي الأساس لإعادة تعريف النوع. الجنين الذكر يكتسب خصائص الجنس الأنثوي. ذكر النموذج النووي ، الذكور الغدد التناسلية ، الأنثى النمط الظاهري. نسب الجسم من الإناث ، وهناك الغدد الثديية ، تقصير المهبل ، والخصيتين - في الشفرين الكبيرين ، القناة الأربية ، تجويف البطن.
قرار من الطابق
نتيجة التعرض للهرمونات أو نتيجة لعلم مستقبلات الخلية المستهدفة ، يمكن إعادة تعريف الجنس يحدث (متلازمة موريس ، إزالة الغدد الجنسية في الديكة).
في الطبيعة ، العديد من العوامل تضعف عمل الجينات التي تتحكم في تطور الجنس. على سبيل المثال ، عند البشر ، في المناسل لشخص واحد ، فإن الخصيتين والمبايض (الخصية والمبيض ينموان) تتطوران على حد سواء خنوثة - ظاهرة ذات صلة بالمساواة بين الجنسين.
استنادا إلى البيانات السريرية ، هناك 3 أنواع من ثنائيي الجنس:
1) الخنوثة الحقيقية: وجود الخلايا الجنسية من كلا الجنسين ؛
2) ذكر كاذب خنثى: هناك خصيتي فقط (الخصيتين ، الخصيتين) ، النمط الظاهري للأنثى ؛
3) pseudohermaphroditism الإناث: لا يوجد سوى المبيضين ، النمط الظاهري الذكور.
هذا التصنيف لا يتطابق مع القواعد الوراثية الخلوية ، حيث أن 46 XY في الرجل هناك خيارات.
نسبة الجنس
يجب أن تكون نسبة الجنس الأساسية (في وقت الإخصاب قريبة من 1: 1 ، حيث أن اجتماع الكروموسومات الجنسية يكون متساوياً). خلال الفحص في البشر ، وجد أن 140-160 من الذكور يتشكلون لكل 100 أنثى لقاح منقوش. Spermatozoa تحتوي على كروموسوم Y أخف وزنا وأكثر حركة ولها شحنة سالبة كبيرة (البيضة لها شحنة موجبة). لذلك ، الحيوانات المنوية التي تحتوي على Y تقوم بتخصيب البويضة في أغلب الأحيان.
التعليم الثانوي - يولد 103-105 ولداً لكل 100 فتاة. قدرة الأمشاج الأنثوية ، البروتينات الأجنبية للأجنة الذكورية. في سن العشرين ، يوجد 100 فتى لكل 100 فتاة.
التعليم العالي - في سن الخمسين ، هناك 85 رجلاً لكل 100 امرأة ، وفي عمر 85 عامًا ، يوجد 50 رجلًا لكل 100 امرأة. الكائن الأنثوي أكثر تكيفًا ، والذي يمكن تفسيره ، إلى جانب أسباب أخرى ، بفسيفساء الكائن الأنثوي على الكروموسومات الجنسية.
فرضية M. Loyon حول الفسيفساء الأنثوية على الكروموسومات الجنسية.
في عام 1949 ، وجد M. Borr و C. Bertrand أن مجموعة من لونين شديد الكثافة وجدت في نواة الخلايا العصبية للنساء. في نوى خلايا الرجال لا تكتشفها. يسمى هذا التجمع باسم الكروماتين الجنسي (جسم بورا) ويمثل كروموسوم X معطل.
في بداية تطور الجنين الأنثوي ، يعمل كل من الكروموسومات X ، أي ضعف عدد الرجال لديهم الجينات. هذا قد يفسر قدرة أكبر للبيضة الملقحة.
في عام 1962 ، وضع ليون فرضية حول تعطيل كروموسوم X واحد في جسم الثدييات الأنثوي. في الجنين الأنثوي ، يعمل كلا الكروموسوم حتى اليوم 16 من التطور الجنيني. في اليوم السادس عشر ، يتم تعطيل كروموسوم واحد مع تشكيل الكروماتين الجنسي. هذه العملية عشوائية ، لذلك فإن حوالي 1/2 من الخلايا النشطة تحافظ على كروموسوم X الأمومي X 5M 0 ، ويتم تعطيل الأب. في حالات أخرى ، يكون الأب نشطًا (X 5O 0) ، ويعطل نشاط الأم. لا يحدث التنشيط. تحتوي كروموسومات X الوالدية والأبوية على جينات أليلية ، ولكنها ليست جينات متطابقة تمامًا ، أي أليل مهيمن يقع في كروموسوم واحد ، والآخر متنحي. امتلاك جينات إضافية يوسع القدرات التكيفية للجسم.
مستويات DIAGRAM الجنسي من الاختلاف الجنسي
التفاضل
┌─┐ ┌┴┐
لعب X └┬┘ └─┘ Y
البويضة الحيوانات المنوية
┌───────────────────┐
الوراثي العضلي الوراثي XX و XY │
└───────────────────┘
┌─────────┴─────────┐
gonadal │ غير معروف متباينة │
g enye الغدد التناسلية من الجنين
│ └───────────────────┘
ant │ H Y مستضد
في الفترة ┌───────┴───────┐ ┐─────────────┐
الأعضاء التناسلية │ │ │ │1 الأعضاء التناسلية │
النضج │ │ │ ┌─│ علامات
│ │ │ │ └────────────┘
in ┌─────┴─────┐┐ ут في الرحم
│ │ غونداد │ │gunad ├────┤ 7-32 أسبوع
│ │ المبيضان │ الخصيتان │ الهرمونات andro
│ └───────────┘ └───────────┘┘ │ │ جينات الخلية
│ │ │ Leydinga
│ │ البلوغ │ │ ┌────────────┐
│ │ التمايز
الهرمونات الهرمونية الغدة النخامية ، الذكور والإناث │ │ العصب العصبي │
│ الهرمونات: الاندروجين (التستوستيرون) ، طرق est est est est est est est
└─ الجينات (progisterone) │ الجنس │
┌──────────────┴───────────────┐ └────────────┘
phenotypic │ الخصائص الجنسية الثانوية признаки
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
النفسية │ الانتماء الجنسي والسلوكي │
└───────────────────────────────────────────────────┘
جسد الأنثى أكثر مقاومة للإشعاع البارد ، والإشعاعات المؤينة ، والحمل الزائد العاطفي (تبكي النساء في كثير من الأحيان ، تبرز الأمينات النشطة بالدموع ، ونتيجة لذلك ينخفض ضغط الدم).
إذا كانت الفرضية تعمل دون قيود ، فلن يكون هناك اختلافات ظاهرية بين النساء الأصحاء مع اثنين من كروموسومات X والمرضى الذين يعانون من X4 0 أو الرجال XY / XXYY. من الواضح أن الكروموسوم X غير معطل بالكامل.
تنظيم نسبة الجنس.
إذا وضعت السائل المنوي في حقل كهربائي ثابت ، فسيكون هناك فصل للكروموسومات X و Y. تستخدم في الماشية. من الممكن الحصول على 80٪ من أحفاد الجنس المرغوب.
الوعي بالانتماء إلى الذكر أو الأنثى هو جزء لا يتجزأ من الإدراك العقلي للجنس ، والبارامترات العقلية. Transsexualism - الخنوثة النفسية. ، ظاهرة انتهاك الهوية الجنسية للفرد. كانت حالات الوعي الوسواسي للأشخاص الذين ينتمون إلى الجنس الآخر معروفة منذ العصور القديمة. لذلك ، وصف هيرودوت ، في سرّه "المرض السكيثي" ، رجلًا لم يرتد ملابس النساء فقط ، بل تبنى صفات الشخصية للجنس الآخر. بعض الأباطرة كان لديهم ميل للتنكر: على سبيل المثال ، كاليجولا. ج. د.
التناقض بين المعلمات التشريحية والعقلية هو نزاع بين البيئة الداخلية والاجتماعية. في جسد الذكر تعيش روح الأنثى ، والعكس بالعكس. قم بإجراء تصحيح جراحي للأرضية. لتحويل الرجل إلى امرأة ، تحتاج إلى عملية واحدة: يتكون المهبل من جلد القضيب وكيس الصفن. من امرأة لجعل الرجل أكثر صعوبة: 3-4 عمليات (إزالة الغدد الثديية ، وتشكيل عضو). هؤلاء الناس محكومون بالعذاب الأبدي: فهم يشربون الهورمونات ، وليس لديهم أطفال. معهد التكاثر البشري وتنظيم الأسرة في موسكو.
التفريق الجنسي ينطوي على تكوين الابتدائي (الغدد التناسلية) والخصائص الجنسية الثانوية.
يتم تحديد الجنس في معظم الحيوانات وراثيا في وقت الإخصاب.
يحمل الكروموسوم X جينات تأنيث الخصية (Tfm) ، طبيعي
يحدد أليله تخليق مستقبل البروتين في الأندروجينات ،
التي يتم توليفها في كل من الكائنات والذكور.
──────┐ - كل كروموسوم هو Tfm غير وظيفي - وهذا يعني
┌┬┬┐ K┌┬┬┐ الغش يطور القشرة.
├──────┐┐ └┴┴┘ ├──────┐┐ XY - يحتوي على الجين I 5HA-gene 0 ، وهو المسؤول عن
│ └──────┘ │ توليف المستضد ، والذي يحدد
│ M │ male = مسؤول عن صنع
│ │ هرمون تستوستيرون.
is ────┴───┐ لوحظ التمايز من الغدد التناسلية في 7-10
│┌┬┬┬┐│ │ │ أسبوع من الحياة. في الأسبوع 10 ، يمكن ممارسة الجنس
│ │ │ │ لتحديد مجموعة من الكروموسومات.
└─────┘ └───────┘
يتم تحديد اتجاه التنمية من خلال وجود كروموسوم Y. عادة ، تحتوي كروموسومات XX على جين معقم يمنع نمو نوع الذكر. هذا التطور يعتمد على مستضد HY (ON) ، الجين I 5HA مسؤول عن إنتاج التستوستيرون. تم العثور عليها في عدة أنواع (وصفها في الفأرة في عام 1955 ، Eychwald ، Silmser). يفرز من قبل الخلايا الجرثومية الذكور الابتدائية.
عادة ، في الأفراد الذين لديهم مجموعة من الكروموسومات الجنسية ، تهيمن الجينات الأنثوية على الجينات التي تحدد الجنس الأنثوي ، ونوع الذكر ، يسيطر الجنس الذكري.
يتم تشكيل الأعضاء التناسلية من مجرى Müller و Wolf ، الذي ينشأ من الكلية الأولية. في النساء ، تتطور قنوات Müller إلى قناة فالوب والرحم ، وضمور أنابيب الذئب. في الذكور ، تتطور قنوات الذئب إلى القنوات المنوية والحويصلات المنوية. يتم توليف هرمون الستيروئيدات التستوستيرون و dihydrotestosterone في الخصيتين الجنينية تحت تأثير هرمون الأم (gonadotropin المشيمي). هذه الهرمونات تعمل على البراعم biopotent.
الأعضاء التناسلية الخارجية والداخلية: قنوات الذئب ، القنوات مولروفي وجيب الجيوب التناسلية. يحدث كائن ذكري طبيعي إذا كانت جميع هذه العناصر تعمل. في غيابها ، يتم تشكيل خصائص الجنس الأنثوي. تطور غير مكتمل من النمط الظاهري الذكوري مع الذكور
النمط الوراثي (الذكورية الكاذبة).
الاستراتيجية التناسلية الحديثة:
- التلقيح الصناعي
- الإخصاب في المختبر.
- النمو الاصطناعي للجنين ونقله إلى الرحم ؛
- الأم البديلة.
معظم الحيوانات هي كائنات ثنائية الجنس. يمكن النظر إلى النوع كمجموعة من السمات والهياكل التي توفر طريقة لإعادة إنتاج النسل ونقل المعلومات الوراثية. يتم تحديد الجنس في معظم الأحيان في وقت الإخصاب ، وهذا هو ، النمط النووي للالميغوت يلعب الدور الرئيسي في تحديد الجنس. يحتوي النمط النووي لكل كائن حي على كروموسومات هي نفسها في كلا الجنسين - الجسيدات الصليبية ، والكروموسومات التي يختلف فيها الجنس الأنثوي والذكور عن بعضها البعض - الكروموسومات الجنسية. في البشر ، الكروموسومات الجنسية "الأنثوية" هي اثنين من الكروموسومات X. أثناء تكوين الأمشاج ، يتلقى كل بويضة واحدة من الكروموزومات X. ويطلق على نوع الجنس الذي يتكون فيه الأمشاج من النوع نفسه الذي يحمل الكروموسوم X اسم homogametic. في البشر ، والجنس الأنثوي هو homogametic. الكروموسومات الجنسية "ذكر" في البشر هي كروموسوم X وكروموسوم Y. أثناء تكوين الأمشاج ، يتلقى نصف الحيوانات المنوية الكروموسوم X ، النصف الآخر من الكروموسوم Y. يسمى الطابق الذي يحتوي على أنواع مختلفة من الأمشاج heterogametic. في البشر ، فإن جنس الذكر هو heterogametic. إذا تم تشكيل زايغوت يحمل اثنين من الكروموزومات X ، فسيتم تكوين جسم أنثوي منه ، إذا كان الكروموسوم X والصبغي Y ذكران.
في الحيوانات ، يمكن تمييز ما يلي أربعة أنواع من تحديد الجنس الكروموسومي.
1. الجنس الأنثوي هو homogametic (XX) ، والجنس هو heterogametic (XY) (الثدييات ، على وجه الخصوص ، والرجل ، ذبابة الفاكهة).
مخطط جيني لتحديد الجنس الصبغية في البشر:
مخطط وراثي لتحديد الجنس الكروموسومي في ذبابة الفاكهة:
2. الجنس الأنثوي هو homogametic (XX) ، جنس الذكر هو heterogametic (X0) (orthoptera).
مخطط جيني لتحديد جنس الكروموسومات في الجراد الصحراوي:
3. الجنس الأنثوي هو متغاير (XY) ، والجنس الذكر هو homogametic (XX) (الطيور والزواحف).
مخطط وراثي لتحديد جنس الكروموسومات في الحمام:
Yandex.DirectAll ads
4. الجنس الأنثوي هو متغاير (X0) ، والجنس الذكر هو homogametic (XX) (بعض أنواع الحشرات).
تحديد الجنس في البشر يحدث بواسطة آلية XY. في نفس الوقت ، الجنس المتغاير هو ذكر ، homogametic - الأنثى. ينقسم تحديد الجندر إلى ثلاث مراحل: الكروموسومات ، الغدد التناسلية والمظهر الظاهري.
قاعدتين أساسيتين لتحديد الجنس في الثدييات
وضعت الدراسات الجنينية الكلاسيكية قاعدتين لتحديد الجنس في الثدييات. تمت صياغة أول هذه في 1960s من قبل ألفريد جوست استنادا إلى التجارب لإزالة جرثومة الغدد التناسلية في المستقبل (أسطوانة الغدد التناسلية) في أجنة الأرانب في وقت مبكر: إزالة بكرات قبل تشكيل الغدد التناسلية أدى إلى تطوير جميع الأجنة مثل الإناث. واقترح أن يتم إفراز الغدد التناسلية للذكور من هورمون التستوستيرون ، المسؤول عن ذكور الأجنة ، وتم توقع وجود مستجيب ثانٍ لهرمون مضاد ل Muller (MIS) ، يتحكم مباشرة في مثل هذه التحولات التشريحية. تم صياغة نتائج الملاحظات كقاعدة: يحدد التخصص في تطوير الغدد التناسلية في الخصية أو المبيض التفريق الجنسي اللاحق للجنين.
حتى عام 1959 ، كان من المفترض أن يكون عدد الكروموسومات X هو العامل الأهم في التحكم بالجنس في الثدييات. ومع ذلك ، فإن اكتشاف الكائنات الحية ذات كروموسوم X واحد ، يتطور كإناث ، والأفراد الذين لديهم كروموسوم Y واحد وكروموسومات X متعددة ، والتي تطورت كذكور ، أجبرتهم على التخلي عن هذه المفاهيم. تم صياغة القاعدة الثانية لتحديد الجنس في الثدييات: يحوي Y-chromosome المعلومات الوراثية المطلوبة لتحديد الجنس عند الذكور.
يسمى أحيانًا الجمع بين القاعدتين السابقتين مبدأ النمو: يحدد الجنس الكروموسومي ، المصاحب لوجود أو عدم وجود كروموسوم Y ، تمايز الغدد التناسلية الجنينية ، والتي بدورها تتحكم في الجنس المظهري للكائن الحي. تسمى هذه الآلية لتحديد الجنس بالوراثة. GSD) ونعارض ذلك ، على أساس الدور المسيطر للعوامل البيئية (م. ESD) أو نسبة الكروموسومات الجنسية و autosomes (eng. CSD).
تحديد الجنس الهرموني
يمكن تمثيل تعريف الجندر كعصا ، والتي تنقلها الآلية الصبغية إلى مناسل غير متمايزة تتطور إلى أجهزة جنسية ذكورية أو أنثى. عند دراسة دور الكروموسومات الجنسية في تطوير الغدد التناسلية ، تبين أن وجود الكروموسوم Y أو غيابه حاسم لدى البشر. في حالة عدم وجود كروموسوم Y ، يحدث تباين التناسلية في المبايض وتتطور المرأة. في وجود كروموسوم Y ، يتطور النظام الذكري. من الواضح أن الكروموسوم Y ينتج مادة تحفز على التمايز الخصوي. "يبدو أن الخطة الأساسية للطبيعة هي جعل المرأة ، وأن إضافة الكروموسوم Y تنتج رجلًا مختلفًا". تستمر المرحلة التالية من التتابع الهرمونات التي تحدد عملية التمايز الجنسي للجنين وتطوره التشريحي. عند الولادة ، ينتهي الجزء الأول من البرنامج. بعد الولادة ، تنتقل العصا إلى العوامل البيئية التي تكمل تشكيل الجنس - عادة ، ولكن ليس دائما ، وفقا للجنس الجيني. تحديد الجنس هو عملية معقدة ومتعددة الخطوات ، والتي تعتمد على البشر ، إلى جانب البيولوجية ، على العوامل النفسية والاجتماعية. هذا يمكن أن يؤدي إلى تغيير الجنس ، أو الجنس الآخر ، أو الجنس الآخر أو السلوك المثلي.
الأساس الفسيولوجي لتحديد الجنس التناسلية
الأساس الفسيولوجي لآلية تحديد الجنس هو ثنائي الجنس من الغدد التناسلية الجنينية في الجنين. في مثل هذه السلالات ، توجد قناة Mullerov وقناة الذئب في وقت واحد - جراثيم القناة التناسلية ، على التوالي ، للإناث والذكور. يبدأ التحديد الأولي للجنس بمظهر خطوط الخلايا المتخصصة في البروجونات ، وهي خلية سيرتولي. في هذا الأخير ، يتم توليف هرمون مضاد للمولر تنبأ به Zhost ، وهو المسؤول عن تثبيط مباشر أو غير مباشر لتطوير قناة مولر - جرثان قناتي فالوب المستقبلية والرحم.
آلية وراثية لتحديد الجنس
كروموسوم Y البشري ، مما يشير إلى موقع الجين SRY
في عام 1987 ، قام ديفيد بيج وزملاؤه بفحص رجل XX الذي ورث جزءًا معينًا من الكروموسوم Y 280 ألف نيوكليوتيد في الطول ، وامرأة XY بحذف يلتقط هذه المنطقة كنتيجة لتبادل المقاطع بين الكروموسومات. توجد هذه القطعة في الكروموسوم Y من جميع حيوانات Eutheria الحقيقية وتقع على مسافة 100 ألف زوج أساسي من حدود جين ZFY pseudoautosomal بطول 140 ألف زوج قاعدي.
تم العثور على ZFY homologue - الجين ZFX على الكروموسوم X ، و ZFX غير معطلة. يقوم العاملان ZFX و ZFY بتشفير عوامل النسخ المحتوية على زخارف إصبع الزنك مع نشاط ربط الحمض النووي. وقد أدى التحليل التفصيلي الإضافي لتسلسلات محددة من الكروموزومات Y لدى الأفراد ذوي الانعكاس الجنسي إلى البحث عن مساحة 35 كيلوبت في الحجم ، وأدى ذلك إلى اكتشاف جين يُعتبر معادلاً حقيقياً للغة الإنجليزية الكلاسيكية. عامل تحديد الخصية. يسمى هذا الجين SRY (eng. تحديد الجنس منطقة Y الجينات).
SRY تقع في مجال تحديد الجنس وتحتوي على مجال محافظ (HMG-box) يشفر بروتينًا من 80 وحدة من الأحماض الأمينية. لوحظ نشاط الجين SRY قبل بداية فترة التمايز للهروب إلى الخصية - 10-12 يوم من تطور الجنين الفأري ، وعلى الأقل في هذه المرحلة ، لا يعتمد على وجود الخلايا الجرثومية. تؤدي الطفرات المحددة أو الحذف في صندوق HMG لهذا الجين في النساء XY إلى انعكاس الجنس. أدى نقل جزء من DNA 14 kbp يحتوي على هذا الجين مع المناطق المرافقة إلى بويضة مخصبة من فرد homogametic باستخدام microinjection في ظهور ذكر مع نمط karyotype XX.
وظائف الجين SRY
إن المجال المشفر بواسطة صندوق HMG لجين SRY يرتبط بشكل خاص بالحامض النووي ويؤدي إلى الانحناء. يمكن أن ينقل ميكانيكيا الانحناء الذي يسببه بروتين SRY أو نظائره التي تحتوي على نطاق HMG على مسافة بعيدة ويلعب دورا هاما في تنظيم النسخ والتكرار وإعادة التركيب. تحتوي منطقة الحمض النووي (DNA) التي يتم فيها ترجمة SRY على جينتين تقوم بترميز الإنزيمات الرئيسية المتضمنة في تمايز الغدد التناسلية الأولية من الذكور: جينات أروماتاز P450 ، التي تتحكم في تحويل التستوستيرون إلى استراديول ومثبط تطور مجرى ميلر ، والذي يسبب نموهما المعاكس ويعزز التمايز الخصية. .
أيضا ، يشارك منتج الجين SRY في عمليات التمايز الجنسي في تفاعل وثيق مع جين آخر ، يدعى الجين Z ، الذي يتكون عادة من وظائفه في تثبيط جينات معينة من الذكور. في حالة النمط الوراثي الذكري العادي 46XY ، يقوم جين SRY بتشفير بروتين يثبط جين Z ، وتنشط جينات معينة من الذكور. في حالة النمط الوراثي الطبيعي للإناث 46XX ، الذي لا توجد فيه SRY ، يتم تنشيط الجين Z ويثبط جينًا محددًا من الذكور ، مما يخلق ظروفًا لتطور النوع الأنثوي.
نظرية الكروموسومات لتحديد الجنس
النظر في تحديد الجنس الكروموسومي. من المعروف أن نسبة الجنس في الكائنات الحيّة ثنائية العين (الحيوانات والنباتات الثنائية) تكون عادة 1: 1 ، أي أن الذكور والإناث شائعة على حد سواء. تتزامن هذه النسبة مع التقسيم في تحليل الصليب ، عندما يكون أحد الأشكال المتقاطعة متغاير الزيجوت (أأ)، والآخر متماثل للأليلات المتنحية (أأ). في ذرية في هذه الحالة هناك تقسيم فيما يتعلق أأ: 1aa. إذا كان الجندر موروثًا وفقًا لنفس المبدأ ، فسيكون من المنطقي الافتراض أن جنسًا واحدًا يجب أن يكون متماثلًا وزوجًا آخر غير متماثل. ثم يجب أن يكون التقسيم حسب الجنس مساوياً لـ 1.1 في كل جيل ، وهو ما يتم ملاحظته بالفعل.
النظرية التطورية لنوع الجنس اقترح جيوداكسيان في عام 1965. وتوضح النظرية من وجهة نظر موحدة العديد من الظواهر المتعلقة بالجنس: الشكلية الجنسية في الصحة والمرض ، ونسبة الجنس ، والوفيات التفاضلية ومعدل الاستجابة الجنسية ، ودور الكروموسومات الجنسية ، والهرمونات الجنسية ، وعدم تناسق الدماغ واليد ، الآثار المتبادلة ، والاختلافات النفسية والاجتماعية بين الجنسين.
تقوم النظرية على مبدأ الأنظمة الفرعية المترافقة التي تتطور بشكل غير متزامن. ذكر هو تشغيل النظام الفرعي للسكان ، الإناث - محافظ النظام الفرعي. تصل المعلومات الجديدة من البيئة لأول مرة إلى الجنس الذكري وفقط بعد انتقال أجيال عديدة إلى الأنثى ، لذلك تطور الجنس الذكري يسبق تطور الأنثى. هذا التحول مرة (اثنان مرحلة تطور سمة) يخلق شكلين من سمة (ذكرا وأنثى) - اختلال الشكل الجنسي في السكان. تعد "المسافة" التطورية بين الأنظمة الفرعية ضرورية للبحث والتحقق من الابتكارات.
 أكثر الأمراض فظاعة في العالم
أكثر الأمراض فظاعة في العالم هيمنة كاملة وغير كاملة
هيمنة كاملة وغير كاملة كيف يمكنك أن تجعل صديقتك سعيدة
كيف يمكنك أن تجعل صديقتك سعيدة