Základy teórie chromozómov. Morganova chromozómová teória: definícia, základy a vlastnosti
Tvorba chromozomálnej teórie v rokoch 1902-1903. Americký cytológ W. Setton a nemecký cytológ a embryológ T. Bo-Veri nezávisle objavili paralelismus v správaní génov a chromozómov počas tvorby gamet a oplodnenia. Tieto pozorovania slúžili ako základ predpokladu, že gény sa nachádzajú v chromozómoch. Experimentálnym dôkazom o lokalizácii špecifických génov v špecifických chromozómoch sa však dostal až v roku 1910 americký genetik T. Morgan, ktorý v nasledujúcich rokoch (1911-1926) dokázal chromozomálnu teóriu dedičnosti. Podľa tejto teórie je prenos dedičných informácií spojený s chromozómami, v ktorých sú gény lokalizované lineárne v určitej sekvencii. Preto sú to chromozómy, ktoré predstavujú materiálny základ dedičnosti.
Tvorbu chromozomálnej teórie uľahčili údaje získané pri štúdiu genetiky pohlavia, keď sa stanovili rozdiely v množine chromozómov v organizmoch rôznych pohlaví.
Križovanie sa vyskytuje v profeze I meiózy počas konjugácie homológnych chromozómov (obrázok 3.10). V tomto čase sa môžu časti týchto dvoch chromozómov pretínať a vymieňať ich časti. V dôsledku toho vznikajú kvalitatívne nové chromozómy, ktoré obsahujú sekcie (gény) materských a otcovských chromozómov. jedinci, ktoré sú získané z takých gamet s novou kombináciou alel, sa nazývajú prekríženie alebo rekombinantné.
Frekvencia (percento) priesečníka medzi dvoma génmi umiestnenými v tom istom chromozóme je úmerná vzdialenosti medzi nimi. Prekročenie medzi týmito dvoma génmi sa vyskytuje menej často, čím sú navzájom bližšie. Vzhľadom k tomu, že vzdialenosť medzi génmi sa zvyšuje, zvyšuje sa pravdepodobnosť, že ich prekročenie rozdelí pozdĺž dvoch rôznych homológnych chromozómov.
Vzdialenosť medzi génmi charakterizuje silu ich uchopenia. Existujú gény s vysokým percentuálnym priľnavosťou a tie, v ktorých je priľnavosť takmer nedetekovateľná. Pri spojení s dedením však maximálna hodnota prekročenia nepresiahne 50%. Ak je vyššia, potom existuje voľná kombinácia medzi dvojicami alel, nerozlíšiteľná od nezávislého dedičstva.
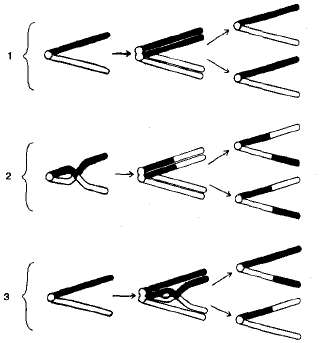
Obr. 3.10. Schéma prekračovania: I - nedostatok prechodu; 2 - prekročenie v štádiu dvoch chromozómov; 3 - prekročenie v štádiu štyroch chromatidov.
Zvážte jeden z prvých pokusov T. Morgan o štúdiu súvisiaceho dedičstva. Pri prechode ovocných much, ktoré sa líšia v dvoch pároch spojených znakov - sivé s jednoduchými krídlami a čierne s normálnymi krídlami - hybridy F 1 boli sivé s normálnymi krídlami (obrázok 3.11).
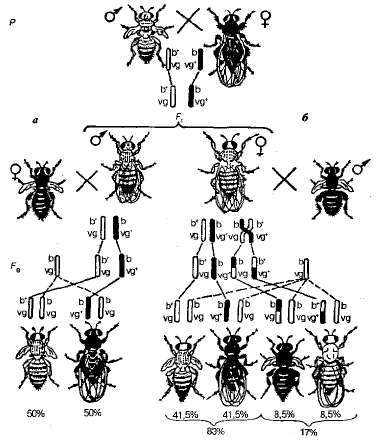
Obr. 3.11, Dedičnosť spojených znakov v Drosophile: a-úplná adhézia (muž bez prechodového digerozozigrtenu); b - krížová spojka (diheterozigattová žena, ktorej kríž nie je potlačený); b +, b - šedá a čierna farba karosérie; vg + , vg - - normálne a rudimentárne krídla.
Ďalej boli vykonané dva typy analytických krížení. V prvom z nich boli odobraté diheterozygotné samce. F 1, a boli prekonané samice homozygotné pre recesívne alely av druhej boli heterosexné samice prekročené s mužmi recesívnymi pre obe znaky (čierne telo a rudimentárne krídla). Výsledky týchto krížení sa ukázali byť odlišné (pozri obrázok 3.11).
V prvom prípade Potomci sa získali potomkovia s rodičovskými (P) rodičovskými fenotypmi pre tento pokus, to znamená šedé muchy s rudimentárnymi krídlami a čiernymi muchami s normálnymi krídlami v pomere 1: 1. Preto táto diheterozygota tvorí len dva druhy gamét. (b + vg a bvg +) namiesto štyroch. Na základe tohto štiepenia možno predpokladať, že muž má kompletnú väzbu génov.
V druhom prípade v F 2 bolo pozorované ďalšie rozdelenie. Okrem rodičovských kombinácií znakov sa objavili nové - letí s čiernym telom a rudimentárnymi krídlami, rovnako ako so šedým telom a normálnymi krídlami. Je pravda, že počet rekombinantných potomkov je malý a predstavuje 17% a riadiace číslo je 83%. Dôvodom pre vznik malého počtu much s novými kombináciami znakov je prekročenie, čo vedie k novej rekombinantnej kombinácii alel génov. b a vg v homológnych chromozómoch. Tieto výmeny sa vyskytujú s pravdepodobnosťou 17% a nakoniec poskytujú dve triedy rekombinantov s rovnakou pravdepodobnosťou - 8,5% každej.
Biologický význam kríženia je extrémne vysoký, pretože genetická rekombinácia umožňuje vytvoriť nové, predtým neexistujúce kombinácie génov a tým zvýšiť dedičnú variabilitu, ktorá poskytuje organizmu dostatok príležitostí na prispôsobenie sa rôznym podmienkam prostredia. Osoba špecificky vykonáva hybridizáciu s cieľom získať potrebné kombinácie možností pre použitie v šľachtiteľskej práci.
Pojem genetickej mapy. T. Morgan a jeho spolupracovníci K. Bridges a A. Stertevanti G. Moler experimentálne ukázali, že znalosť fenoménu väzby a prekračovania umožňuje nielen vytvorenie spojovacej skupiny génov, ale aj vytvorenie genetických máp chromozómov, ktoré indikujú poradie umiestnenia génov v chromozóme a relatívnom vzdialenosť medzi nimi.
Mapa genetických chromozómov nazýva schému vzájomného usporiadania génov v rovnakej spojovacej skupine. Takéto mapy sa zostavujú pre každý pár homológnych chromozómov.
Možnosť takéhoto mapovania je založená na stálosti percentuálneho prekročenia medzi určitými génmi. Genetické mapy chromozómov sa zostavujú pre mnohé druhy organizmov: hmyz (Drosophila, komáre, šváby atď.), Huby (kvasinky, aspergilus), baktérie a vírusy.
Prítomnosť genetickej mapy naznačuje vysoký stupeň štúdia jedného alebo iného typu organizmu a má veľký vedecký záujem. Takýto organizmus je výborným predmetom ďalšej experimentálnej práce, ktorá má nielen vedecký, ale aj praktický význam. Najmä znalosť genetických máp vám umožňuje naplánovať prácu na získavaní organizmov s určitými kombináciami znakov, ktoré sa v súčasnosti široko používajú v chovateľskej praxi. Teda vytvorenie kmeňov mikroorganizmov schopných syntetizovať proteíny, hormóny a iné komplexné organické látky potrebné pre farmakológiu a poľnohospodárstvo je možné len na základe metód genetického inžinierstva, ktoré sú založené na poznatkoch genetických máp príslušných mikroorganizmov.
Ľudské genetické mapy môžu byť tiež užitočné v zdravotníctve a medicíne. Poznatky o lokalizácii génu v konkrétnom chromozóme sa používajú pri diagnostike radu závažných dedičných ľudských ochorení. Už existuje príležitosť na génovú terapiu, t.j. na opravu štruktúry alebo funkcie génov.
Porovnávanie genetických máp rôznych druhov živých organizmov tiež prispieva k pochopeniu evolučného procesu.
Hlavné ustanovenia chromozomálnej teórie dedičnosti. Analýza javov súvisiaceho dedičstva, prekračovanie, porovnanie genetických a cytologických máp nám umožňuje formulovať hlavné ustanovenia chromozomálnej teórie dedičnosti:
- Gény sú lokalizované v chromozómoch. Zároveň rôzne chromozómy obsahujú rôzne počty génov. Okrem toho je súbor génov každého nehomologického chromozómu jedinečný.
- Alelové gény obsadzujú rovnaké lokusy v homológnych chromozómoch.
- Gény sa nachádzajú v chromozóme v lineárnej sekvencii.
- Gény toho istého chromozómu tvoria väzbovú skupinu, vďaka ktorej dochádza ku zjednotenému dedičstvu určitých znakov. V tomto prípade je sila adhézie nepriamo spojená so vzdialenosťou medzi génmi.
- Každý druh je charakterizovaný špecifickým súborom chromozómov - karyotypu.
Zdroj : NA Lemeza L. V. Kamluk N.D. Lisov "Príručka biológie pre vstup do vysokých škôl"
Práca Morgana položila základy chromozomálnej teórie dedičnosti, ukázali, že obmedzenia voľnej kombinatoriky niektorých génov sú spôsobené umiestnením týchto génov v jednom chromozóme a ich fyzickou väzbou.
Morgan zistil, že väzba génov nachádzajúcich sa na tom istom chromozóme nie je absolútna. Počas meiózy môžu chromozómy jedného páru vymieňať homológne miesta medzi sebou pomocou procesu, ktorý sa nazýva prekríženie. Čím ďalej od seba sú gény v chromozóme, tým častejšie sú zdieľané prechodom. Na základe tohto javu sa navrhlo miera adhézie génov - percento prekročenia - a vytvorili sa prvé genetické mapy chromozómov pre rôzne druhy Drosophila.
Odroda Drosophila bola vybraná ako objekt genetickej analýzy a Morgan študovala dedičstvo rôznych vlastností.
Prekročenie homozygotnej samice s šedým telom a dlhými krídlami (dominutie) s homozygotným krátkokrídlým mužom s čiernymi krídlami v F1 - jednotnosť (sivé telo, dlhé krídla)
Ukázalo sa, že výsledky budú rôzne v závislosti od pohlavia hybridu.
Ak muž bol hybrid, potom potomstvo produkovalo 2 fenotypové triedy, ktoré úplne opakovali charakteristiky rodičov.
Ak bola samica hybridom, potom sa v nerovnom poradí ukázalo 4 fenoménové triedy potomkov. Väčšina potomstva (83%) je potomkom s rodičovskými črtami, menej (17%) sú jedinci s novými kombináciami znakov.
Morgan dospela k záveru, že spojka môže byť neúplná, kde sa skupina spojky pretína prekrížením.
Neobvyklá percento u potomkov vzhľadom k tomu, že crossover nie je vždy crossover frekvencia závisí od vzdialenosti medzi génmi - čím väčšia je vzdialenosť, tým nižšia je pevnosť spoja medzi génmi, tým viac crossoveru.
Gamety, ktoré majú chromozómy, ktoré neboli prekrížené, sa nazývajú non-crossover.
Ak sa v gaméti chromozómy, ktoré prešli cez kríženie, sú kríženie.
6. Hlavné ustanovenia chromozomálnej teórie dedičnosti
1. Gény sa nachádzajú v chromozómoch lineárne v určitých oblastiach - lokusoch. Alelové gény obsadzujú rovnaké lokusy homológnych chromozómov.
2. Gény nachádzajúce sa v tom istom chromozóme tvoria väzbovú skupinu a sú zdedené alebo spojené. Počet spojovacích skupín = počet chromozómov v haploidnej množine.
3. Križovanie je možné medzi homológnymi chromozómami, čím sa zlomí spojenie
4. proces prechodu je priamo úmerný vzdialenosti medzi génmi.
Prekročenie o 1% = 1 ° C
7. Koncepcia cytoplazmatickej dedičnosti
Prítomnosť určitého množstva dedičného materiálu v cytoplazme v podobe kruhových molekúl DNA mitochondrií a plastidov, a iné špecifické-jadrových genetických prvkov vedie špeciálne k zastaveniu ich účasť na tvorbe fenotypu v procese individuálneho rozvoja.
Cytoplazmatické gény nepodliehajú mendelským zákonom dedičnosti, ktoré sú určené chovaním chromozómov v mitóze, meióze a oplodnení. Vzhľadom na to, že organizmus vytvorený v dôsledku hnojenia dostáva cytoplazmatické štruktúry hlavne s vajíčkovými bunkami, cytoplazmatická dedičnosť znakov sa uskutočňuje cez materskú líniu. Tento druh dedičstva bol prvýkrát opísaný v roku 1908 K. Corrensom, pokiaľ ide o charakteristiku pestrých listov v niektorých rastlinách.
A hnojenie. Tieto pozorovania slúžili ako základ predpokladu, že gény sa nachádzajú v chromozómoch. Experimentálne dôkazy o lokalizácii špecifických génov v špecifických chromozómoch sa však získali iba v meste amerického genetika T. Morgana, ktorý v nasledujúcich rokoch (-) potvrdil chromozomálnu teóriu dedičnosti. Podľa tejto teórie je prenos dedičných informácií spojený s chromozómami, v ktorých sú gény lokalizované lineárne v určitej sekvencii. Preto sú to chromozómy, ktoré predstavujú materiálny základ dedičnosti.
Tvorbu chromozomálnej teórie uľahčili údaje získané pri štúdiu genetiky pohlavia, keď sa stanovili rozdiely v množine chromozómov v organizmoch rôznych pohlaví.
Genetika podlahy
Podobný spôsob určenia pohlavia (typu XY) je vlastný všetkým cicavcom, vrátane ľudí, ktorých bunky obsahujú 44 autozómov a dva X chromozómy u žien alebo XY chromozómy u mužov.
To znamená, Stanovenie pohlavia typu XY, alebo typ ovocného muštu a človeka, - najbežnejší spôsob určenia pohlaviacharakteristické pre väčšinu stavovcov a niektorých bezstavovcov. Typ X0 sa vyskytuje vo väčšine ortopérov, ploštice, chrobákov, pavúkov, v ktorých chromozóm Y neexistuje, takže muž má genotyp X0 a ženskú XX.
Vo všetkých vtákoch, väčšina motýľov a niektorých plazov, samci sú homogametickí a ženy sú heterogametické (typu XY alebo typu XO). Pohlavné chromozómy týchto druhov sú označené písmenami Z a W, aby sa izolovala táto metóda určovania pohlavia; súbor chromozómov mužov je označený symbolom ZZ a samice symbolom ZW alebo Z0.
Dôkaz, že pohlavné chromozómy určujú pohlavie organizmu, bol získaný pri štúdiu nedisjunkcie pohlavných chromozómov v Drosophile. Ak jeden z gamét získať dva pohlavné chromozómy, a druhá - nikoho, že zlúčenie týchto pohlavných buniek s normálnym môže premeniť individuálne sadou pohlavných chromozómov XXX, XO, XXY, atď. Bolo zistené, že v Drosophila jednotlivcov so sadou HO. - Psy , a so súborom HHU - samíc (u ľudí - naopak). Jednotlivci so súborom XXX majú hypertrofované znaky samice (super-samice). (Jedinci so všetkými týmito chromozomálnymi odchýlkami sú sterilné v Drosophile). Neskôr sa ukázalo, že v Drosophile je pohlavie určované pomerom (rovnováha) medzi počtom X chromozómov a počtom súborov autozómov.
Dedičnosť pohlavných znakov
V prípade, že gény, ktoré riadia tvorbu konkrétneho znaku, sú lokalizované v autozómoch, dedenie sa vykonáva bez ohľadu na to, ktorý rodič (matka alebo otec) je nosičom študovaného znaku. Ak sú gény v pohlavných chromozómoch, dramaticky sa mení charakter dedičnosti postáv. Napríklad v Drosophile, gény lokalizované na X chromozóme, spravidla nemajú alely na chromozóme Y. Z tohto dôvodu sa recesívne gény v chromozóme X heterogametického pohlavia takmer vždy objavujú a sú v jedinečnom.
Znaky, ktorých gény sa nachádzajú na pohlavných chromozómoch, sa nazývajú pohlavne viazané znaky. Fenomén pohlavne viazaného dedičstva objavil T. Morgan v spoločnosti Drosophila.
U ľudí majú chromozómy X a Y homológnu (pseudoautozomálnu) oblasť, kde sa nachádzajú gény, ktorých dedičstvo sa nelíši od génov autozomálnych.
Okrem homologických oblastí majú chromozómy X a Y aj nehomologické oblasti. Nehomologická oblasť chromozómu Y okrem génov, ktoré určujú mužské pohlavie, obsahuje gény membrán medzi prstami a chlpaté uši u ľudí. Patologické príznaky spojené s nehomologickou oblasťou chromozómu Y sa prenášajú na všetkých synov, pretože od otca dostávajú Y chromozóm.
Nehomologická oblasť chromozómu X obsahuje množstvo génov, ktoré sú dôležité pre životne dôležitú aktivitu organizmov. Vzhľadom na to, že heterogamatózny sex (XY) X chromozómu je reprezentovaný v singulári, znaky určené génmi nehomologickej oblasti chromozómu X sa objavia, aj keď sú recesívne. Tento stav génov sa nazýva hemizygotný. Príkladom takýchto X-viazaných recesívnych znakov u ľudí je hemofília, svalová dystrofia Duchenne, atrofia optického nervu, farebná slepota (farebná slepota) atď.
Hemofília je dedičné ochorenie, pri ktorom krv stráca schopnosť zrážať sa. Rana, dokonca aj poškriabaniu alebo modrínovaniu, môže spôsobiť veľké vonkajšie alebo vnútorné krvácanie, ktoré často vedie k smrti. Toto ochorenie sa zriedkavo vyskytuje iba u mužov. Zistilo sa, že obidve najbežnejšie formy hemofílie (hemofília A a hemofília B) sú spôsobené recesívnymi génmi umiestnenými na chromozóme X. Ženy (nosiče), ktoré sú heterozygotné pre tieto gény, majú normálne alebo mierne znížené zrážanie krvi.
Fenotypový prejav hemofílie u dievčat sa bude pozorovať, ak je matka dievčaťa nosičom hemofílie a otec je hemofilik. Podobný model dedičnosti je charakteristický aj pre iné recesívne pohlavné znaky.
Prepojené dedičstvo
Nezávislá kombinácia vlastností (tretí zákon Mendel) sa uskutočňuje pod podmienkou, že gény, ktoré určujú tieto vlastnosti, sú v rôznych pároch homológnych chromozómov. Preto v každom organizme je počet génov, ktoré sa môžu nezávisle kombinovať v meióze, obmedzený počtom chromozómov. V tele však počet génov výrazne prevyšuje počet chromozómov. Napríklad viac ako 500 génov bolo skúmaných v kukurici pred obdobím molekulárnej biológie, viac ako 1 000 u drosophila múch a asi 2 000 u ľudí, zatiaľ čo 10, 4 a 23 párov chromozómov. Skutočnosť, že počet génov vo vyšších organizmoch dosahuje niekoľko tisíc, bol pre W. Suttona jasný už na začiatku 20. storočia. To naznačuje, že mnohé chromozómy sú lokalizované na každom chromozóme. Gény lokalizované v rovnakom chromozóme tvoria väzbovú skupinu a zdedia spolu.
Spoločné dedičstvo génov T. Morgan navrhlo nazvať súvisiace dedičstvo. Počet spojovacích skupín zodpovedá haploidnému počtu chromozómov, pretože spojovacia skupina pozostáva z dvoch homológnych chromozómov, v ktorých sú umiestnené rovnaké gény. (U jedincov heterogamického pohlavia, napríklad u mužov cicavcov, skupiny spojok sú v skutočnosti ešte jedna, pretože chromozómy X a Y obsahujú rôzne gény a sú to dve odlišné spojovacie skupiny, takže ženy majú 23 spojovacích skupín a pre mužov - 24).
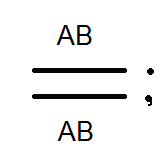
Spôsob dedenia spojených génov sa líši od dedičnosti génov lokalizovaných v rôznych pároch homológnych chromozómov. Takže ak nezávislá kombinácia diheterozygotná osoba tvorí v rovnakých množstvách štyri typy gamét (AB, Ab, aB a ab), potom s reťazcovým dedičstvom (v prípade absencie prekríženia) tvorí rovnaký digerozygot iba dva typy gamét: (AB a ab) rovnaké množstvá. Posledný z nich opakuje kombináciu génov v chromozóme rodičov.
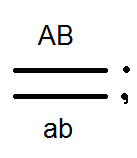
Zistilo sa však, že okrem bežných (bez boxov) gamét existujú iné (krížové) gaméty s novými kombináciami génov - Ab a aB, ktoré sa líšia od kombinácií génov v chromozómoch rodičov. Príčinou takýchto gamét je výmena častí homológnych chromozómov alebo prechod.
Križovatka sa vyskytuje v profeze I meiózy počas konjugácie homológnych chromozómov. V tomto čase sa môžu časti týchto dvoch chromozómov pretínať a vymieňať ich časti. V dôsledku toho vznikajú kvalitatívne nové chromozómy, ktoré obsahujú sekcie (gény) materských a otcovských chromozómov. Jednotlivci, ktorí sú získaní z takýchto gamét s novou kombináciou alel, sa nazývajú kríženie alebo rekombinantné.
Frekvencia (percento) priesečníka medzi dvoma génmi umiestnenými v tom istom chromozóme je úmerná vzdialenosti medzi nimi. Prekročenie medzi týmito dvoma génmi sa vyskytuje menej často, čím sú navzájom bližšie. Vzhľadom k tomu, že vzdialenosť medzi génmi sa zvyšuje, zvyšuje sa pravdepodobnosť, že ich prekročenie rozdelí pozdĺž dvoch rôznych homológnych chromozómov.
Vzdialenosť medzi génmi charakterizuje silu ich uchopenia. Existujú gény s vysokým percentuálnym priľnavosťou a tie, v ktorých je priľnavosť takmer nedetekovateľná. Pri spojení s dedením však maximálna frekvencia prekročenia nepresiahne 50%. Ak je vyššia, potom existuje voľná kombinácia medzi dvojicami alel, nerozlíšiteľná od nezávislého dedičstva.
Biologický význam kríženia je extrémne vysoký, pretože genetická rekombinácia umožňuje vytvoriť nové, predtým neexistujúce kombinácie génov a tým zvýšiť dedičnú variabilitu, ktorá poskytuje organizmu dostatok príležitostí na prispôsobenie sa rôznym podmienkam prostredia. Osoba špecificky vykonáva hybridizáciu s cieľom získať potrebné kombinácie možností pre použitie v šľachtiteľskej práci.
Pojem genetickej mapy
T. Morgan a jeho spolupracovníci K. Bridges, A. G. Sturtevant a G. J. Meller experimentálne ukázali, že znalosť fenoménu spojenia a prekračovania umožňuje nielen vytvorenie väzbovej skupiny génov, ale aj vytvorenie genetických máp chromozómov, na ktorých je uvedený poradie lokalizácie. génov v chromozóme a relatívnych vzdialenostiach medzi nimi.
Genetická mapa chromozómov sa nazýva schéma vzájomného usporiadania génov, ktoré sú v rovnakej spojovacej skupine. Takéto mapy sa zostavujú pre každý pár homológnych chromozómov.
Možnosť takéhoto mapovania je založená na stálosti percentuálneho prekročenia medzi určitými génmi. Genetické mapy chromozómov sa zostavujú pre mnohé druhy organizmov: hmyz (Drosophila, komáre, šváby atď.), Huby (kvasinky, aspergilus), baktérie a vírusy.
Prítomnosť genetickej mapy naznačuje vysoký stupeň štúdia jedného alebo iného typu organizmu a má veľký vedecký záujem. Takýto organizmus je výborným predmetom ďalšej experimentálnej práce, ktorá má nielen vedecký, ale aj praktický význam. Najmä znalosť genetických máp vám umožňuje naplánovať prácu na získavaní organizmov s určitými kombináciami znakov, ktoré sa v súčasnosti široko používajú v chovateľskej praxi. Teda vytvorenie kmeňov mikroorganizmov schopných syntetizovať proteíny, hormóny a iné komplexné organické látky potrebné pre farmakológiu a poľnohospodárstvo je možné len na základe metód genetického inžinierstva, ktoré sú založené na poznatkoch genetických máp príslušných mikroorganizmov.
Ľudské genetické mapy môžu byť tiež užitočné v zdravotníctve a medicíne. Poznatky o lokalizácii génu v konkrétnom chromozóme sa používajú pri diagnostike radu závažných dedičných ľudských ochorení. Už existuje príležitosť na génovú terapiu, ktorou je korekcia štruktúry alebo funkcie génov.
Hlavné ustanovenia chromozomálnej teórie dedičnosti
Analýza javov súvisiaceho dedičstva, prekračovanie, porovnanie genetických a cytologických máp nám umožňuje formulovať hlavné ustanovenia chromozomálnej teórie dedičnosti:
- Gény sú lokalizované v chromozómoch. Zároveň rôzne chromozómy obsahujú rôzne počty génov. Okrem toho je súbor génov každého nehomologického chromozómu jedinečný.
- Alelové gény obsadzujú rovnaké lokusy v homológnych chromozómoch.
- Gény sa nachádzajú v chromozóme v lineárnej sekvencii.
- Gény toho istého chromozómu tvoria väzbovú skupinu, to znamená, že sú zdedené prevažne spojené (spoločne), vďaka čomu sú niektoré znaky spojené s dedičstvom. Počet spojovacích skupín sa rovná haploidnému počtu chromozómov daného druhu (homogametického pohlavia) alebo viac o 1 (v heterogametickom pohlaví).
- Priľnavosť je rozbitá v dôsledku prechodu, ktorého frekvencia je priamo úmerná vzdialenosti medzi génmi v chromozóme (preto je sila adhézie nepriamo spojená so vzdialenosťou medzi génmi).
- Každý druh je charakterizovaný špecifickým súborom chromozómov - karyotypu.
zdroje
- N. A. Lemeza L. V. Kamlyuk N. D. Lisov "Príručka o biológii pre vstup na univerzity"
poznámky
Nadácia Wikimedia. 2010.
Tvorcom teórie chromozómov (HT) je vedec Thomas Morgan. CT je výsledkom štúdie dedičnosti na bunkovej úrovni.
Podstata teórie chromozómov:
Materiálom nosičov dedičnosti sú chromozómy.
Hlavným dôkazom toho je:
Cytogenetický paralelizmus
Stanovenie pohlavia chromozómov
Pohlavne viazané dedičstvo
Spojenie génov a prechod
Hlavné ustanovenia teórie chrosomal:
Dedičinové sklony (gény) sú lokalizované v chromozómoch.
Gény sa nachádzajú v chromozóme lineárnym spôsobom.
Každý gén zaberá špecifickú oblasť (lokus). Alelové gény obsadzujú podobné lokusy v homológnych chromozómoch.
Gény lokalizované v jednom chromozóme sú zdedené spolu, prepojené (Morganov zákon) a tvoria spojovaciu skupinu. Počet skupín spojok sa rovná počtu haploidných chromozómov (n).
Medzi homológnymi chromozómmi možné výmenné miesta alebo rekombinácia.
Vzdialenosť medzi génmi sa meria v percentách prekročenia - morganov.
Frekvencia prekračovania je nepriamo úmerná vzdialenosti medzi génmi a súdržná sila medzi génmi je nepriamo úmerná vzdialenosti medzi nimi.
Cytogenetický paralelizmus
Odborník z Morgan Sytton poznamenal, že správanie génov podľa Mendela sa zhoduje s chovaním chromozómov: (Tabuľka - cytogenetický paralelizmus)
Každý organizmus nesie 2 dedičné ložiská, gamét zahŕňa len 1-v dedičnom ložisku páru. Pri oplodnení v zygote a neskôr v tele, 2 dedičné zálohy na každom základe.
Chromozómy sa správajú rovnakým spôsobom, čo naznačuje, že gény ležia v chromozómoch a zdedia spolu s nimi.
Stanovenie pohlavia chromozómov
V roku 1917 Allen ukázal, že muži a ženy v mechoch sa líšia v súbore chromozómov. V bunkách diploidného tkaniva mužského tela, pohlavných chromozómov XiY, u ženského XiX. Chromozómy teda definujú takéto označenie ako pohlavie a preto môžu byť materiálnymi nositeľmi dedičnosti. Neskôr sa ukázalo, že chromozómové stanovenie pohlavia pre iné organizmy, vrátane ľudí. (Tabuľka)
Pohlavne viazané dedičstvo
Vzhľadom na to, že pohlavné chromozómy sú odlišné v mužských a ženských organizmoch, znaky, ktorých gény sa nachádzajú v chromozómoch X alebo Y, budú dedične odlišné. Takéto označenia sa nazývajú pohlavných znakov.
Vlastnosti dedičnosti pohlavných znakov
Nepozorovali 1 Mendelov zákon
Recipročné križovatky poskytujú rôzne výsledky.
Existuje krížový kríž (alebo dedičstvo krížom).
Po prvýkrát dedičstvo spojené s týmto rysom objavil Morgan v Drosophile.
|
W + červené oči |
(C) x w + x w + x x w y |
(C) x w x w * x w + y |
|||||
|
w - biele oči | |||||||
|
(CC) X W + X w - červené oči |
X w X W + - červené oči |
||||||
|
(CM) X W + Y - červené oči |
X w Y - Biele oči |
||||||
|
Takže dedičnosť mutácie identifikovanej Morganom - "biele oči" - biela, bola charakterizovaná znakmi uvedenými vyššie: Zákon jednotnosti nebol dodržaný Na 2 vzájomných prechodoch dostali rôzne potomky Na druhom priechode majú synovia znamenie matky (biele oči), dcéry - znamenie otca (červené oči). Tento druh dedičstva sa nazýva krížové dedičstvo. |
|||||||
(TABLE dedičstvo viazané na pohlavie)
Pohlavne viazané dedičstvo je spôsobené absenciou génov v chromozóme Y, ktoré sú alelické pre gény chromozómov X. Y chromozóm je omnoho menší ako chromozóm X, v súčasnosti je 78 lokalizovaných v ňom (?) génov, zatiaľ čo vo svojom chromozóme viac ako 1098.
Príklady dedičnosti súvisiacej s pohlavím:
Hemofília, dystrofia Duchenne, Duncanov syndróm, Alportov syndróm atď.
Existujú gény, ktoré sú naopak obsiahnuté v chromozóme Y a chýbajú v chromozóme X, a preto sa nachádzajú iba v mužských organizmoch a nikdy nie u žien (holandské dedičstvo) a prenášajú sa iba na synov od otca.
Spojenie génov a prechod
V genetike bol takýto fenomén ako "príťažlivosť génov" známy: niektoré nealérové vlastnosti neboli zdedené nezávisle, ako Mendelov zákon by mal robiť, ale zdedili spolu, nedali nové kombinácie. Morgan vysvetlil to tým, že tieto gény sú v tom istom chromozóme, takže sa rozchádzajú do dcérskych buniek spoločne v jednej skupine, akoby boli spojené. Tento fenomén nazval - združené dedičstvo.
Morgan Clutch Law:
Gény nachádzajúce sa v tom istom chromozóme sa dedia spolu, spojené.
Gény umiestnené v tom istom chromozóme tvoria väzbovú skupinu. Počet skupín spojok sa rovná "n" - haploidnému počtu chromozómov.
Homozygotné línie muchov so šedou farbou trupu a dlhými krídlami a muchami s čiernym telom a krátkymi krídlami boli prekročené. Gény farby tela a dĺžky krídel sú spojené, t.j. ležia v jednom chromozóme.
|
A - sivé telo čierne telo B- normálne krídla (dlhé) b - základné krídla |
(CF) AABBxaabb (CM) |
|||||
|
Šedá dlho okrídlená |
Čierna krátkosrstá |
|||||
|
Zaznamenajte v chromozomálnom vyjadrení |
||||||
|
Šedé telo Dlhé krídla |
Čierne telo Krátke telo |
|||||
|
|
|
|||||
|
Všetky mušky majú šedé telo a dlhé krídla. |
||||||
|
tj v tomto prípade sa dodržiava zákon o jednotnosti hybridov I generácie. Avšak vo F 2 namiesto očakávaného rozdelenia 9: 3: 3: 1 bol získaný pomer 3 sivých dlhých krídel k 1 časti čiernych krátkych krídel; Nové kombinácie znakov sa nezobrazili. Morgan navrhol, že deheterozygote F2 - ( |
||||||
|
|
||||||
|
Šedé telo Dlhé krídla |
Čierne telo Krátke telo |
|||||
|
|
|
|||||
|
F |
|
|
||||
|
Šedé telo Dlhé krídla |
Čierne telo Krátke krídla |
|||||
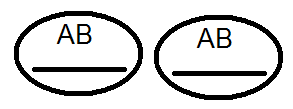
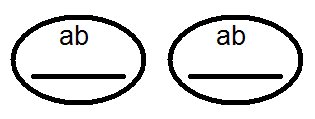
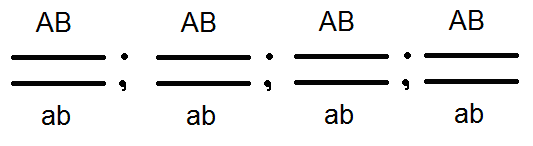
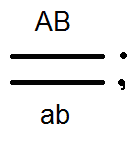 ) produkujú (dať) gamét nie 4, ale iba 2 typy -
) produkujú (dať) gamét nie 4, ale iba 2 typy -  a
a  , Analýza vykonaných kríží potvrdila toto:
, Analýza vykonaných kríží potvrdila toto: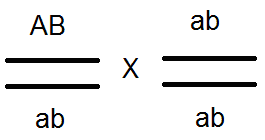
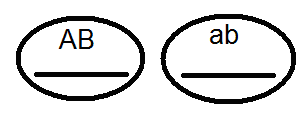
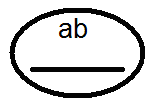
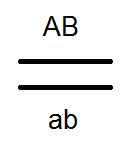
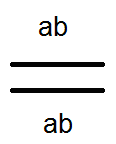
V dôsledku toho dochádza k rozdeľovaniu vo F 2, ako pri monohybridnom prekročení 3: 1.
|
|
||||
|
|
|
|||
|
|
|
|
|
|
|
Šedé telo Dlhé krídla |
Šedé telo Dlhé krídla |
Šedé telo Dlhé krídla |
Čierne telo Krátke krídla |
|
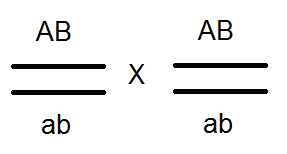
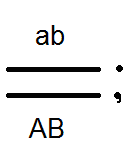
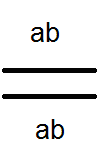
Prekračovanie.
V malej percento prípadov vo F 2 v pokusoch Morgana sa objavili muchy s novými kombináciami znakov: krídla sú dlhé, telo je čierne; krídla sú krátke a telo je šedé. tj Známky "odomknuté." Morgan vysvetlil to tým, že chromozómy počas konjugácie na génoch výmeny meiózy. V dôsledku toho sa jednotlivci získajú s novými kombináciami znakov, t.j. ako sa hodí tretieho zákona Mendela. Morgan nazval túto rekombináciu génovej výmeny.
Neskôr cytológovia skutočne potvrdili hypotézu Morgana, ktorá našla výmenu chromozómových oblastí v kukurici a salámandroch. Tento proces nazývali krížom.
Prekračovanie zvyšuje rozmanitosť potomstva v populácii.
Prečo deti vyzerajú ako ich rodičia? Prečo sú niektoré choroby bežné v niektorých rodinách, ako je napríklad daltonizmus, polydaktyly, hypermobilita kĺbov, cystická fibróza? Prečo existuje séria ochorení, s ktorými sú choré iba ženy a iné sú len muži? Dnes všetci vieme, že odpovede na tieto otázky je potrebné hľadať v dedičnosti, teda v chromozómoch, ktoré dieťa dostáva od každého z rodičov. A moderná veda vďačí týmto poznatkom Thomasovi Huntovi Morganovi - americkej genetike. Popísal proces prenosu genetických informácií a spolu s kolegami vyvinul chromozomálnu teóriu dedičnosti (často nazývanú Morganovu chromozomálnu teóriu), ktorá sa stala základným kameňom modernej genetiky.
História objavovania
Bolo by nesprávne povedať, že Thomas Morgan bol prvý, kto sa zaujímal o prenos genetických informácií. Prví výskumníci, ktorí sa pokúsili porozumieť úlohe chromozómov v dedičstve, sa môžu považovať za diela Chistyakov, Beneden, Rabelya v 70 - 80 rokoch 19. storočia.
Potom neboli žiadne mikroskopy tak silné, aby ste mohli vidieť chromozomálne štruktúry. A samotný termín "chromozóm" nebol. V roku 1888 ho predstavil nemecký vedec Heinrich Valdeyr.
Nemecký biológ Theodor Bowery v dôsledku svojich experimentov dokázal, že pre normálny vývoj tela potrebuje pre svoj druh normálny počet chromozómov a ich nadbytok alebo nedostatok vedie k závažným vývojovým chybám. Postupom času bola jeho teória skvelo potvrdená. Možno povedať, že chromozomálna teória T. Morgana získala svoj východiskový bod práve preto, že Boveriho výskum.
Začiatok výskumu
Zovšeobecniť existujúce vedomosti o teórii dedičnosti, Thomas Morgan ich dokázal doplniť a rozvinúť. Ako objekt pre svoje pokusy si vybral ovocnú mušku a nie náhodou. Bol to ideálny objekt pre štúdium prenosu genetických informácií - len štyri chromozómy, plodnosť, krátka životnosť. Morgan začal výskum pomocou čistých línií muchov. Čoskoro zistil, že v zárodočných bunkách je jedna sada chromozómov, to znamená 2 miesto 4. Morgan určil ženský pohlavný chromozóm ako X a mužský ako Y.
Pohlavne viazané dedičstvo
Morganova chromozomálna teória ukázala, že existujú určité sexuálne spojené črty. Predná zraka, s ktorou vedecký pracovník vykonával svoje pokusy, normálne má červenú farbu očí, ale v dôsledku mutácie tohto génu sa v populácii objavili biele-oči a medzi nimi bolo oveľa viac mužov. Gén, ktorý je zodpovedný za farbu očí múch, je lokalizovaný na X chromozóme, nie je na chromozóme Y. To znamená, že keď je žena prekrížená, na jednom chromozóme X, ktorý je mutovaným génom, a muž s bielym očkom, pravdepodobnosť prítomnosti tejto vlastnosti v potomstve bude mať vzťah k pohlaviu. Najjednoduchší spôsob zobrazenia na diagrame:
- P: XX "x X" Y;
- F 1: XX, XY, X "X", X "Y.
X - pohlavný chromozóm samice alebo muža bez génu bielych očí; X "- chromozóm s génom bielych očí.
Dešifrujeme výsledky kríženia:
- XX "- žena s červenými očami, nosičom génu bývalých očí.Vďaka prítomnosti druhého X chromozómu sa tento mutovaný gén" prekrýva "zdravým a vo fenotype sa tento rytmus nezobrazuje.
- X "Y je muž s bielym očkom, ktorý dostal od matky chromozóm X s mutovaným génom. Vzhľadom na prítomnosť iba jedného X chromozómu nemá mutantný znak nič, čo by zablokoval a zdá sa, že sa vyskytuje vo fenotype.
- X "X" - biela-eyed žena, zdedil pozdĺž chromozómu s mutantný gén od matky a otca. U samice, iba ak obidve X chromozómy nesú gén bielych očí, objaví sa vo fenotype.
Thomas Morgana vysvetlil mechanizmus dedičnosti mnohých genetických ochorení. Keďže na chromozóme X existuje viac génov než na chromozóme Y, je jasné, že je zodpovedný za väčšinu vlastností organizmu. X chromozóm z matky sa prenáša na synov a dcéry spolu s génmi zodpovednými za vlastnosti tela, vonkajšie príznaky, choroby. Spolu s X-linked je dedičstvo spojené s Y. Ale U-chromozóm je iba u mužov, takže ak sa v ňom vyskytne nejaká mutácia, môže sa vysielať iba mužským potomkom.
Morganova chromozomálna teória dedičnosti pomohla porozumieť modelu prenosu genetických ochorení, ale ťažkosti spojené s ich liečbou sa doteraz nevyriešili.
![]()
Prekračovanie
V priebehu výskumu objavil fenomén kríženia študent Thomas Morgan, Alfred Stertevant. Ako ukazujú ďalšie experimenty, vďaka prekračovaniu sa objavujú nové kombinácie génov. Je to ten, kto porušuje proces reťazového dedičstva.
Takže chromozomálna teória T. Morgany získala ďalšiu dôležitú pozíciu - medzi tým je prechod a jej frekvencia je určovaná vzdialenosťou medzi génmi.
Hlavné ustanovenia
Na systematizáciu výsledkov experimentov vedcov sme prezentovali hlavné body chromozomálnej teórie Morgan:
- Znaky tela závisia od génov stanovených v chromozómoch.
- Gény jedného chromozómu sú prenášané na potomstvo spojené. Pevnosť takéhoto spojenia je väčšia, tým menšia je vzdialenosť medzi génmi.
- V homológnych chromozómoch sa pozoruje fenomén kríženia.
- Keď poznáme frekvenciu prekročenia určitého chromozómu, dá sa vypočítať vzdialenosť medzi génmi.

Druhý bod Morganovej chromozómovej teórie sa tiež nazýva Morgan Rule.
uznanie
Výsledky výskumu boli vnímané brilantne. Morganova chromozomálna teória bola prelomom v biológii dvadsiateho storočia. V roku 1933 bol vedec ocenený Nobelovou cenou za objav úlohy chromozómov v dedičnosti.
O niekoľko rokov neskôr dostal Thomas Morgan medailu Copley za dokonalosť v genetike.
Teraz sa v škole študuje Morganova chromozomálna teória dedičnosti. Venovala veľa článkov a kníh.
Príklady dedičstva súvisiaceho s pohlavím
Morganova chromozomálna teória ukázala, že vlastnosti organizmu sú determinované génmi, ktoré sú v ňom stanovené. Základné výsledky, ktoré prijal Thomas Morgan, poskytli odpoveď na otázku týkajúcu sa prenosu takých chorôb ako je hemofília, Low syndróm, farebná slepota, Bruttonova choroba.
Ukázalo sa, že gény všetkých týchto ochorení sú umiestnené na chromozóme X a u žien tieto choroby vyskytujú oveľa menej často, pretože zdravý chromozóm môže blokovať chromozóm génom choroby. Ženy, ktoré to nevedia, môžu byť nositeľmi genetických ochorení, ktoré sa potom prejavujú u detí.
U mužov sa X-viazané ochorenia alebo fenotypové príznaky prejavujú, pretože neexistujú žiadne zdravé X chromozómy.
T. Morganova chromozomálna teória dedičnosti sa používa pri analýze rodinnej anamnézy pre genetické choroby.
 Vták s červeným pruhom na hlave
Vták s červeným pruhom na hlave Nové znamenie zverokruhu - Ophiuchus
Nové znamenie zverokruhu - Ophiuchus Neobvyklé fenomény prírody
Neobvyklé fenomény prírody