Ženské pohlavie je vždy určené kombináciou xx. Podlahová biológia
Genetika podlahy
Pohlavie je charakterizované komplexom znakov určených génmi umiestnenými v chromozómoch. V bunkách ľudského tela sú chromozómy spárované diploidné súbory. U druhov s dvojdomnými jedincami nie je chromozomálny komplex mužov a žien rovnaký a líši sa jedným párom chromozómov (pohlavných chromozómov). Rovnaké chromozómy tohto páru boli nazývané X (X) -chromozóm, nepárový, chýbajúci od druhého pohlavia - Y (play) -chromozóm; ostatné, pre ktoré nie sú rozdiely, sú autozómy (A).
Ženské bunky obsahujú dva identické pohlavné chromozómy, ktoré sú označené ako XX, u mužov sú reprezentované dvoma nepárovými chromozómami X a Y. Súbor chromozómov muža a ženy sa teda líši len v jednom chromozóme: chromozómová skupina ženy obsahuje 44 autozómov + XX, mužov - 44 autozómov + XY.
Počas rozdelenia a dozrievania zárodočných buniek u ľudí sa vytvárajú gaméty s haploidným počtom chromozómov: vajcia spravidla obsahujú 22 + X chromozómy. Preto sa u žien tvorí len jeden druh gaméty (gaméty s chromozómom X). U samcov obsahujú gaméty 22 + X alebo 22 + Y chromozómy a tvoria sa dva typy gamét (gamét s chromozómom X a gamétou s chromozómom Y). Ak v priebehu oplodnenia spermie s chromozómom X vstúpi do vajíčka, vytvoria sa zárodočné klíčky a s chromozómom Y - mužským pohlavím.
Preto stanovenie pohlavia osoby závisí od prítomnosti zárodočných buniek muža - spermií hnojením vajíčok, X alebo Y chromozómov.
Existujú štyri hlavné typy chromozómového sexuálneho stanovenia:
1. Mužský pohlavie je heterogametický; 50% génov nesie X-, 50% -U - chromozóm, napríklad človek, cicavce, dipterány, chrobáky, chyby (Snímka 4).
2. Mužský pohlavie je heterogametický; 50% gamét nesie X-, 50% nemá žiadny sexuálny chromozóm, napríklad kobylky, klokany (Snímka 7).
3. Ženský pohlavie je heterogametický; 50% génov nesie X-, 50% gamétový -Y- chromozóm, napríklad vtáky, plazy, obojživelníky, morušová (Snímka 7).
4. Ženský pohlavie je heterogametický; 50% gamét nesie X-, 50% nemá sexuálny chromozóm, napríklad mol.
Dedičnosť znakov, ktorých gény sú lokalizované v pohlavných chromozómoch, sa nazýva dedičstvo, spojený s podlahou.
26. Genotyp ako integrálny systém. Interakcia génov, viacnásobné pôsobenie génov.
Genotyp ako kompletný systém
Vlastnosti génov. Na základe pri pohľade s príkladmi dedičnosti znakov v monochromatických a di-hybridných krížikoch, môže človek nadobudnúť dojem, že genotyp organizmu sa skladá zo súhrnu samostatných, nezávisle pôsobiacich génov, z ktorých každý určuje vývoj svojho vlastného znaku alebo vlastností. Takáto myšlienka priameho a jednoznačného spojenia génu s vlastnosťou často nie je pravdivá. V skutočnosti existuje obrovský počet znakov a vlastností živých organizmov, ktoré sú určené dvomi alebo viacerými pármi génov a naopak, jeden gén často kontroluje mnoho znakov. Okrem toho môže byť vplyv génu zmenený blízkosťou iných génov a okolitých podmienok. Preto v ontogenéze pôsobia nie jednotlivé gény, ale celý genotyp ako integrálny systém so zložitými spojeniami a interakciami medzi jeho zložkami. Tento systém je dynamický: vznik nových alel alebo génov v dôsledku mutácií, tvorba nových chromozómov a dokonca aj nových genómov vedie k pozoruhodnej zmene genotypu v čase.
Povaha prejavu génového účinku v zložení genotypu ako systému sa môže líšiť v rôznych situáciách a pod vplyv rôznych faktorov. To možno ľahko zistiť, ak vezmeme do úvahy vlastnosti génov a zvláštnosti ich prejavu v znakoch:
Gén je vo svojom účinku diskrétny, to znamená, že je izolovaný vo svojej aktivite od iných génov.
Gén je špecifický vo svojom prejave, t.j. je zodpovedný za striktne definovanú charakteristiku alebo vlastnosť organizmu.
Gén môže pôsobiť postupne, t.j. zvyšuje stupeň prejavu znaku s nárastom počtu dominantných alel (dávka génu).
Jeden gén môže ovplyvniť vývoj rôznych znakov - to je viacnásobný alebo pleiotropický účinok génu.
Rôzne gény môžu mať rovnaký vplyv na vývoj rovnakého znaku (často kvantitatívne znaky) - ide o viac génov alebo o polygény.
Gén môže interagovať s inými génmi, čo vedie k vzniku nových príznakov. Takáto interakcia sa uskutočňuje nepriamo - prostredníctvom produktov ich reakcií syntetizovaných pod ich kontrolou.
Účinok génu sa môže modifikovať zmenou jeho polohy na chromozóme (pozitívny účinok) alebo pôsobením rôznych environmentálnych faktorov.
Interakcie alelických génov. Tento fenomén, keď niektoré gény (alely) sú zodpovedné za jeden znak, sa nazýva génová interakcia. Ak sú to alely rovnakého génu, potom sa takéto interakcie nazývajú alelické, a v prípade alel rôznych génov - non-alelické.
Uvádzajú sa tieto hlavné typy alelických interakcií: dominancia, neúplná dominancia, naddominatosť a dominancia.
nadvláda - typ interakcie dvoch alel jedného génu, keď jeden z nich úplne eliminuje manifestáciu druhého. Tento jav je možný za nasledujúcich podmienok: 1) dominantná alela v heterozygotnom stave umožňuje syntézu produktov dostatočných na prejavenie znaku rovnakej kvality ako v stave dominantného homozygotu v materskej forme; 2) recesívna alela je úplne neaktívna alebo produkty jej aktivity neinteragujú s produktmi aktivity dominantnej alely.
Príklady tejto interakcie alelických génov sú dominancia fialovej farba hrachové kvety na bielej farbe, hladké semená nad pokrčenými, tmavé vlasy cez svetlo, hnedé oči po modrej u ľudí atď.
Neúplné dominantné postavenie alebo stredný charakter dedičstva pozorované v prípade, keď sa fenotyp hybridného (heterozygotného) líši od fenotypu oboch rodičovských homozygotov, to znamená, že vyjadrením znaku je medziprodukt s viac alebo menej odchýlkami voči jednému alebo druhému rodičovi. Mechanizmus tohto javu je, že recesívna alela je neaktívna a stupeň aktivity dominantnej alely je nedostatočný na zabezpečenie požadovanej úrovne prejavu dominantného znaku.
Príklad neúplnej dominancie je dedičstvo. farba kvety v rastlinách nočnej krásy (obrázok 3.5). Ako je zrejmé z diagramu, homozygotné rastliny sú buď červené (AA) buď biely (AA) kvety a heterozygotné (AA) - ružová. Pri prechode rastlín s červenými kvetmi a rastlinami s bielymi kvetmi vo F 1, všetky rastliny majú ružové kvety, to znamená, že sú stredný charakter dedičstva. Pri prekračovaní hybridov s ružové kvety v F 2 existuje zhoda štiepenia fenotypom a genotypom, pretože dominantný homozygot (AA) odlišné od heterozygotov (AA). Takže v tomto príklade s nočnými kozmetickými rastlinami, štiepenie v F 2 Farba kvetu je zvyčajne nasledovná - 1 červená (AA): 2 ružový (Aa): 1 biela (AA).
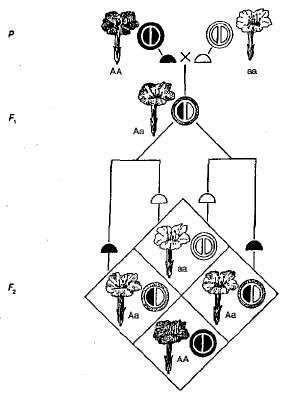
Obr. 3.5. Dedičstvo farby kvetu v prípade neúplnej dominancie nočnej krásy.
Neúplné dominantné postavenie sa rozšírilo. Pozoruje sa v dedičstve vlasov prechádzať u ľudí, farba hovädzieho dobytka, farba peria u kurčiat a mnoho ďalších morfologických a fyziologických znakov rastlín, zvierat a ľudí.
superdominancie - silnejší prejav znaku u heterozygotného jedinca (Aa), než akýkoľvek homozygot (AA a aa). Predpokladá sa, že tento jav je základom heterológie (pozri bod 3.7).
Kodaminirvanie- účasť obidvoch alel na určení znaku u heterozygotného jedinca. Výrazným a dobre skúmaným príkladom dominancie je dedičnosť krvnej skupiny IV u ľudí (skupina AB).
Červené krvinky ľudí v tejto skupine majú dva typy antigénov: antigén (určený genómom dostupným v jednom z chromozómov) a antigénom (určená genómom / a, lokalizovaným na inom homológnom chromozóme). Len v tomto prípade obe alely - 1 (V homozygotná stavová kontrola krvnej skupiny II, skupiny A) a ja B (v homozygotnom stave kontroluje krvnú skupinu III, skupina B). alely 1 a ja B pracujú v heterozygote, ako keby boli navzájom nezávisle.
Príklad dedičnosti skupiny krv ilustruje a prejav viacnásobná alelizmus: gén / môžu byť zastúpené troma rôznymi alelami a existujú gény, ktoré majú desiatky alel. Všetky alely toho istého génu sú pomenované série viacerých alel, z ktorých každý diploidný organizmus môže mať akékoľvek dve alely (a len). Medzi týmito alelami sú všetky uvedené varianty alelických interakcií možné.
Fenomén mnohonásobnej allelizmu je bežný. Existujú rozsiahle série viacerých alel, ktoré určujú typ zlučiteľnosti s hnojením v hubách, opeľovanie v semenných rastlinách, určovanie farby zvieracích chlpov atď.
Interakcie nealylových génov Interakcie nealylových génov sú opísané v mnohých rastlinách a zvieratách. Vedú k objaveniu digerozygotov u potomstva nezvyčajného štiepenia podľa fenotypu: 9: 3: 4; 9: 6: 1; 13: 3; 12: 3: 1; 15: 1, t.j. modifikácie všeobecného Mendelovho vzorca 9: 3: 3: 1. Existujú prípady interakcie dvoch, troch a viacerých nealetických génov. Medzi ne patria tieto hlavné typy: komplementarita, epistáza a polyméry.
komplementárne, alebo ďalšie, Táto interakcia sa nazýva nealícne dominantné gény, v dôsledku čoho sa zjavuje znak, ktorý v oboch rodičoch chýba. Napríklad, keď dve odrôd sladkého hrášku prechádzajú s bielymi kvetmi, potomstvo sa objaví s fialovými kvetmi. Ak označujete genotyp jednej odrody aabb, a druhý aABB,
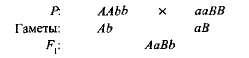
Hybridná prvá generácia s dvoma dominantnými génmi (A a v) získal biochemický základ na výrobu anthocyanínového fialového pigmentu, zatiaľ čo buď jeden gén a Ani jeden z génov B neposkytol syntézu tohto pigmentu. Syntéza anthokyanínu je komplexný reťazec postupných biochemických reakcií kontrolovaných niekoľkými nealetickými génmi a len vtedy, ak existujú najmenej dva dominantné gény (A-B) fialová farba sa vyvíja. V iných prípadoch (aaV- a A-bb) Kvety rastliny sú biele (znamienko "-" vo vzorci genotypu znamená, že toto miesto môže byť obsadené dominantnou aj recesívnou alelou).
Pri samoopelivých rastlinách sladkého hrášku F 1 v F 2 rozdelenie na fialové a biele kvitnúce formy bolo pozorované v pomere blízky 9: 7. Nachádzajú sa fialové kvety 9/1 6 rastlín, biela - na 7/16. Pennetova mriežka jasne ukazuje príčinu tohto javu (obrázok 3.6).
epistáze - ide o typ interakcie génov, v ktorej alely jedného génu potláčajú prejav alelického páru iného génu. gény inhibuje pôsobenie iných génov epistatických inhibítorov alebo potláčajúce. Potlačený gén sa nazýva hypostatické.
Podľa zmeny v počte a pomere fenotypu a ciccov počas dihybridného štiepenia F 2 zvážte niekoľko typov epistatických interakcií: dominantnú epistázu (A\u003e B alebo B\u003e A) s rozdelením 12: 3: 1; recesívna epistáza (a\u003e B alebo b \u003e A), ktorý je vyjadrený rozdelením 9: 3: 4 atď.
polymerism čo sa prejavuje tým, že sa vytvára jedno znamenie vplyv niekoľko génov s rovnakým fenotypovým výrazom. Tieto gény sa nazývajú polymér. V tomto prípade bola prijatá zásada jednoznačného pôsobenia génov na vývoj znaku. Napríklad pri prekročení vrecka pastiera s trojuholníkovým a oválnym plodom vo F 1 sa vytvárajú rastliny s ovocím trojuholníkového tvaru. S ich seba-opelenie v F 2 rozdelenie na rastliny s trojuholníkovým a oválnym luskom v pomere 15: 1. Vysvetľuje to skutočnosť, že existujú dva gény, ktoré konajú jedinečne. V týchto prípadoch sú označené rovnako. 1 a 2 .
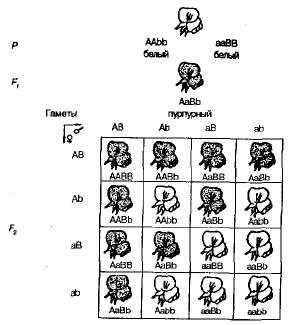
Obr. 3.6 , Dedičstvo farby kvetov v sladkom hrášku
Potom všetky genotypy (A 1 ,- 2 ,-, 1 s 2 a 2 , je 1 1 2 -) bude mať rovnaký fenotyp - trojuholníkové struky a len rastliny a 1 a 1 a 2 2 budú odlišné - tvoria oválne lusky. To je prípad nekumulačných polymérov.
Polymérne gény môžu pôsobiť podľa typu kumulatívnych polymérov. Čím viac podobných génov v genotype organizmu je silnejší prejav tejto vlastnosti, t.j. so zvýšením dávky génu (A 1 2 3 atď.) sa jeho činnosť zhrňuje alebo kumuluje. Napríklad intenzita sfarbenia endospermu pšeničného zrna je úmerná počtu dominantných alel rôznych génov na trihybridnom krížení. Najviac farebné zrná boli 1 1 2 2 3 , A 3 a zrno a 1 a 1 a 2 2 a 3 a 3 nemal pigment.
Podľa typu kumulatívnych polymérov sa zdedia mnohé znaky: mliečnosť, výroba vajec, hmotnosť a iné znaky hospodárskych zvierat; veľa dôležitých parametrov fyzickej sily, zdravia a duševných schopností; dĺžka ucha zŕn; - obsah cukru v koreňoch cukrovej repy alebo v tukoch slnečnicové semená a tak ďalej
Preto početné pozorovania naznačujú, že prejav väčšiny znakov je výsledkom vplyvu komplexu interagujúcich génov a environmentálnych podmienok na formovanie každého špecifického znaku.
Interakcia s génom
Vzťah medzi génmi a vlastnosťami je dosť komplikovaný. V organizme nie vždy jeden gén určuje iba jeden znak a naopak jeden znak je určený len jedným génom. Častejšie môže jeden gén prispieť k prejavu viacerých znakov naraz a naopak. Genotyp organizmu nemožno považovať za jednoduchý súčet nezávislých génov, z ktorých každý funguje mimo kontakt s ostatnými. Fenotypové prejavy znaku sú výsledkom interakcie mnohých génov.
Viacnásobné pôsobenie génov (pleiotropia) - procesy vplyvu jediného génu na tvorbu viacerých znakov.
Napríklad u človeka gén, ktorý určuje farbu červených vlasov, spôsobuje ľahšiu pokožku a pihy.
Niekedy gény, ktoré určujú morfologické charakteristiky, ovplyvňujú fyziologické funkcie, znižujú vitalitu a plodnosť alebo sú smrteľné. Takže gén, ktorý spôsobuje modrú farbu v norke, znižuje plodnosť. Dominantný gén sivého sfarbenia v ovce arakul v homozygotnom stave je podrobný, pretože takéto jahňatá majú nevyvinutý žalúdok a pri prepnutí na trávu umierajú.
Komplementárna génová interakcia. Niekoľko génov môže ovplyvniť vývoj jediného znaku. Interakcia niekoľkých nealetických génov vedúcich k vývoju jediného znaku sa nazýva komplementárna. Napríklad kurčatá majú štyri formy hrebeňa, prejav akéhokoľvek z nich je spojený s interakciou dvoch párov nealetických génov. Rohovitý hrebeň je dôsledkom pôsobenia dominantného génu jednej alely, hrebeňa v tvare hrachu - dominantného génu druhej alely. V hybridoch sa v prítomnosti dvoch dominantných nealkoholových génov vytvorí ostrou-hrebeňovitý hrebeň a za neprítomnosti všetkých dominantných génov, t.j. v recesívnych homozygotiach pre dva neelektrické gény sa vytvorí jednoduchý hrebeň.
Výsledkom interakcie génov je farba vlny u psov, myší, koní, tvaru tekvice, farby sladkých hrachových kvetov.
Polymér je interakcia nealetických génov, keď stupeň vývoja znaku závisí od celkového počtu dominantných génov. Podľa tohto princípu sa zdedí sfarbenie ovsa a pšenice a farba pleti človeka. Napríklad v čiernych farbách existujú 4 dominantné gény v dvoch pároch nealetických génov a žiadny u ľudí s bielou kožou, všetky gény sú recesívne. Kombinácia rôzneho počtu dominantných a recesívnych génov vedie k tvorbe mulatóz s rôznou intenzitou farby kože: od tmavého až po svetlo.
Existujú dve hlavné skupiny génových interakcií: interakcia medzi alelickými génmi a interakciou medzi nealelickými génmi. Treba však pochopiť, že to nie je fyzická interakcia samotných génov, ale interakcia primárnych a sekundárnych produktov, ktoré spôsobia jeden alebo druhý znak. V cytoplazme dochádza k interakcii medzi proteínmi - enzýmami, ktorých syntézu určujú gény alebo medzi látkami, ktoré sa tvoria pod vplyvom týchto enzýmov.
Nasledujúce typy interakcií sú možné:
1) na vytvorenie konkrétneho znaku je potrebná interakcia dvoch enzýmov, ktorých syntéza je určená dvomi nealylovými génmi;
2) enzým, ktorý bol syntetizovaný za účasti jedného génu úplne potláča alebo inaktivuje pôsobenie enzýmu, ktorý bol tvorený iným nealelickým génom;
3) dva enzýmy, ktorých tvorba je riadená dvomi nealylovými génmi, ktoré ovplyvňujú jeden znak alebo jeden proces, takže ich kombinované pôsobenie vedie k vzniku a zvýšeniu prejavu znaku.
Interakcia s allelickým génom
Gény, ktoré zaberajú rovnaké (homológne) lokusy v homológnych chromozómoch sa nazývajú alelické. Každý organizmus má dva alelické gény.
Takéto formy interakcie medzi alelickými génmi sú známe: úplné dominantné postavenie, neúplné dominantné postavenie, dominancia a naddominancia.
Hlavnou formou interakcie je úplná dominancia, ktorú prvýkrát popísal G. Mendel. Jej podstatou je skutočnosť, že v heterozygotnom organizme dominuje manifestácia jednej z alel nad prejavom druhého. S úplnou dominanciou štiepenia genotypom 1: 2: 1 sa nezhoduje s rozdelením na fenotyp - 3: 1. V lekárskej praxi sa z dvoch tisíc monogénnych dedičných chorôb takmer na polovicu vyskytujú dominantné prejavy patologických génov. U heterozygotov sa patologická alela vo väčšine prípadov prejavuje príznakmi ochorenia (dominantný fenotyp).
Neúplná dominancia je forma interakcie, pri ktorej v heterozygotnom organizme (Aa) dominantný gén (A) úplne potláča recesívny gén (a), v dôsledku čoho sa objaví medzipodnik medzi rodičovskými znakmi. Tu je štiepenie genotypu a fenotypu rovnaké a je 1: 2: 1
Keď kodominant v heterozygotných organizmoch, každý alelický gén spôsobuje tvorbu produktu závislého na ňom, to znamená, že produkty obidvoch alel sa ukážu byť. Klasickým príkladom takého prejavu je systém krvných skupín, najmä systém ABO, keď ľudské erytrocyty nesú na povrchu antigény, ktoré sú kontrolované oboma alelami. Táto forma prejavu sa nazýva kodominancia.
Superdominancia - ak je dominantný gén výraznejší v heterozygotnom stave než v homozygotnom stave. Tak v Drosophile, s genotypom AA, normálna dĺžka života; Aa - predĺžený trivatický život; aa - smrteľný.
Viaceré slabosti
Každý organizmus má len dva alelické gény. Často však v prírode môže byť počet alel viac ako dva, ak niektoré lokus môže byť v rôznych štátoch. V takýchto prípadoch sa uvádzajú viaceré alely alebo viacnásobné alleomorfizmy.
Viaceré alely sú označené rovnakým písmenom s rôznymi indexmi, napríklad: A, A1, A3 ... Alely gény sa nachádzajú v rovnakých oblastiach homológnych chromozómov. Pretože vždy existujú dva homológne chromozómy v karyotype, aj s viacnásobnými alelami, každý organizmus môže mať naraz iba dve identické alebo odlišné alely. Len jeden z nich vstupuje do reprodukčnej bunky (spolu s rozdielom v homológnych chromozómoch). Pre viacnásobné alely je charakteristický účinok všetkých alel na ten istý znak. Rozdiel medzi nimi spočíva iba v stupni vývoja znaku.
Druhým znakom je skutočnosť, že somatické bunky alebo bunky diploidných organizmov obsahujú maximálne dve alely niekoľkých, pretože sú umiestnené v rovnakom chromozómovom lokuse.
Ďalšia funkcia je vlastná viacerým alel. Podľa povahy dominancie sú alelomorfné znaky umiestnené v postupnom rade: častejšie normálny nezmenený znak dominuje ostatným, druhý gén série je recesívny vzhľadom na prvý, ale dominuje nasledujúcim atď. Jedným príkladom manifestácie viacerých alel u osoby sú krvné skupiny systému ABO.
Viacnásobná allelis má dôležitý biologický a praktický význam, pretože zvyšuje kombinačnú variabilitu, najmä genotypovú.
Interakcia génov ne-alel
Existuje veľa prípadov, keď charakter alebo vlastnosti sú determinované dvoma alebo viacerými neoddeliteľnými génmi, ktoré navzájom spolupracujú. Hoci tu je interakcia podmienená, pretože to nie sú gény, ktoré interagujú, ale produkty, ktoré kontrolujú. V tomto prípade existuje odchýlka od Mendelovských štiepnych vzorov.
Existujú štyri hlavné typy interakcií génov: komplementarita, epistáza, polyméry a modifikácia (pleiotropia).
Komplementarita je takýto typ interakcie nealetických génov, keď jeden dominantný gén dopĺňa účinok ďalšieho nelélického dominantného génu a spoločne definujú nový znak, ktorý rodičom chýba. Okrem toho sa zodpovedajúca vlastnosť vyvíja iba za prítomnosti obidvoch neallických génov. Napríklad síra je farba vlny u myší kontrolovaných dvoma génmi (A a B). Gene A určuje syntézu pigmentov, avšak homozygóty (AA) a heterozygoty (Aa) sú albínové. Ďalší gén B poskytuje zhromažďovanie pigmentov hlavne na báze a na koncoch vlasov. Kríženie diheterozygotov (AaBb x AaBb) vedie k rozštiepeniu hybridov v pomere 9: 3: 4. Číselné pomery pre komplementárne interakcie môžu byť rovnaké ako 9: 7; 9: 6: 1 (modifikácia štiepenia Mendeleeva).
Príkladom komplementárnej interakcie génov u ľudí môže byť syntéza proteínového proteínu, interferónu. Jeho tvorba v tele je spojená s komplementárnou interakciou dvoch nealelických génov nachádzajúcich sa v rôznych chromozómoch.
Epistáza je interakcia nealetických génov, v ktorých jeden gén inhibuje účinok iného nealeálneho génu. Dominantné aj recesívne gény môžu spôsobiť útlak (A\u003e B, A\u003e B, B\u003e A, B\u003e A) av závislosti od toho epistáza je dominantná a recesívna. Drvivý gén sa nazýva inhibítor alebo supresor. Inhibičné gény všeobecne neurčujú vývoj určitého znaku, ale iba inhibujú pôsobenie iného génu.
Gén, ktorého účinok sa potláča, sa nazýva hypostatická. Pri interakcii s epistotálnymi génmi je štiepenie fenotypu v F2 13: 3; 12: 3: 1 alebo 9: 3: 4, atď. Farba dyňového ovocia, farba koní sú determinované týmto typom interakcie.
Paul - ide o súbor morfologických, fyziologických, biochemických, behaviorálnych a iných znakov tela, ktoré spôsobujú reprodukciu.
Znaky, ktoré sa líšia jednotlivcami rôznych pohlaví, sú rozdelené na primárne a sekundárne. Primárne orgány sú orgány, ktoré zabezpečujú tvorbu gamét a oplodnenie (pohlavné orgány, pohlavné orgány, orgány). Ide o vonkajšie a vnútorné pohlavné orgány, ktoré sa nachádzajú v embryogenéze. Sekundárne - nepodieľajte sa na sexuálnej reprodukcii. Vyvíjajú sa pod vplyvom pohlavných hormónov a objavujú sa v puberte (u ľudí vo veku 12-15 rokov). Sú to znaky vývoja muskuloskeletálneho systému, subkutánneho mastného tkaniva, vlasov, hlasového tónu, správania; vtákoch, spievaní, periach atď.
Príznaky jednotlivcov spojených so sexom sa dajú rozdeliť do troch kategórií:
1) obmedzené na podlahu
2) podlahové riadenie
3) spojený s podlahou.
Vývoj prvého je spôsobený génmi nachádzajúcimi sa v autozómoch obidvoch pohlaví, ale prejavujúc sa len v jednom pohlaví. Takže gény, ktoré tvoria vajíčka, sú prítomné u kurčiat a kohútikov, ale vyskytujú sa iba u kurčiat. Podobne sa správajú gény tuku a výťažku mlieka u hovädzieho dobytka. Tento jav sa pozoruje v súvislosti s expozíciou príslušným pohlavným hormónom.
Príkladom druhej je prejav rohatosti u samíc kráv, komolost - u žien. U ľudí: plešatosť u mužov, dna - 80% u mužov a 12% u žien.
Príznaky, ktorých vývoj je riadený génmi pohlavných chromozómov, sa nazývajú pohlavne viazané. Existuje asi 200 takýchto príznakov. S chromozómom X je spojená farebná slepota, hemofília; s Y-chromozómom - hypertrichóza, ichtyóza.
TYPY STANOVENIA PODLAHY.
1. Progamová - pred oplodnením. Pomer pohlavných chromozómov v tomto prípade nehral úlohu, pretože diploidných oocytov. (Niektoré červy, rotifers - ženy vyvíjajú z veľkých oocytov, samce z malých oocytov).
2. Singamické - genetické stanovenie pohlavia počas oplodnenia, ktoré závisí od povahy kombinácie pohlavných chromozómov alebo pomeru pohlavných chromozómov a autozómov.
3. Epigamické - pod vplyvom vonkajšieho prostredia (červy bonelia).
CHROMOSOMÁLNA DEFINÍCIA PODLABY
┌─────────────────────────┬───────────────────┬──────────────────┐
│ Chromozomálne typy │ genotypy │ druhy gamét │
│ určenie pohlavia ├───────────────────────────────────────────────────────────
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
│ samčia heterogéma │ │
│ Čertský hmyz │ │ │ │
│ (chyby Protenor, chrobáky, │ │ │ │ │
│pauki, kobylky) │ ХО │ ХХ │ Х, О │ Х │
│ Drosophilus │ XY │ XX │ X, Y X │
Ÿ stavovcov │ │ │ │ │
│ (cicavce, človek) │ XY │ XX │ X, Y │ X │
│ │ │ │ │ │
│ samica heterogamu │ │
│ Vtáky. ryby, motýle, │ │ │ │ │
│ kóp │ │ │ │
│ vodné. │ XX │ XY │ X │ X, Y │
Cena a iné bezmocné │ │ │ │ │
Noc │ XX │ XO │ X │ X, O
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
Stanovenie chamozomálneho pohlavia s genetickou kontrolou sa vzťahuje na syngamickú definíciu. Zodpovedá za pohlavie chromozómu, ktorý sa nazýva pohlavie. Normálna mužská gaméta nesie buď X alebo Y chromozóm, a všetky vajcia - X chromozóm. V prípade normálneho rozdielu chromozómov počas meiózy sa tvoria normálne vajíčka a spermie s obvyklým súborom chromozómov X a Y. Pohlavie zygoty je určené pomerom gamét XX a XY (homogametické a heterogametické).
Corrensova chromozomálna teória pohlavia (1907) spočíva v tom, že pohlavie je určované kombináciou pohlavných chromozómov počas oplodnenia. Nasledujúce typy chromozómového stanovenia pohlavia sú rozlíšené: XY, XO, ZW, ZO.
V prípade porušenia priebehu mitózy alebo meiózy sa môžu vytvoriť ginandomorfy. Obsah pohlavných chromozómov v rôznych bunkách týchto jedincov môže byť odlišný (mozaika). V Drosophila lietať: XX a XO, u človeka XX a XY, v súvislosti s ktorými rôzne bolesti tela môžu mať zodpovedajúce príznaky pohlavia. Môžu existovať ďalšie prípady mozaiky: XX / XXX, XY / XXX; XO / XXY a ďalšie
Ak sa pohlavné chromozómy nerozlomia, v ľudských zygátoch môže byť 12 možných kombinácií pohlavných chromozómov, čo je príčinou chromozomálnych aberácií u ľudí.
│ X │ XX │ O
─────┼────────┼─────────┼────────
X │ XX │ XXX │ XO
Y │ XY │ XXY │ YO
XY │ XXY │ XXXY │ XYO
O │ XO │ │
V prípade nondisjunkcie pohlavných chromozómov počas meiózy sa géma XX a O tvoria u žien. a XY a O u mužov. S ich účasťou na oplodnení sa vytvárajú zygoty s nezvyčajnou kombináciou pohlavných chromozómov. U ľudí sa takéto anomálie vyskytujú 1 u 600-700 novorodencov. Zygote Yo zomrie v ranom štádiu; osoby XXX, XXY, XO sú životaschopné a ich pohlavie závisí od prítomnosti alebo neprítomnosti chromozómu "Y", ktorý s akýmkoľvek počtom X chromozómov riadi tvorbu znakov mužského pohlavia, rozvoj a stimuluje tvorbu semenníkov. Prebytok X chromozómov spôsobuje ústavné anomálie a poruchy inteligencie. Ale v prírode existujú jedinci, v ktorých je chromozóm "Y" geneticky inertný a nemá osobitný vplyv na určenie pohlavia.
Vzorky Drosophily typu CW, ktoré boli samce ale neplodné (1916, C. Bridges), boli zistené a osoby XXY sú normálne úrodné ženy.
Teória rovnováhy pohlaví (Bridges, 1922). Študoval pomer pohlavia a automobilov.
U normálnych žien s sadu chromozómov 2n pomer autosome a X-chromozóm je 1: 2n = 2A + 2X (2X: 2A = 1 - normálny žena), 1, 5 - cez žien: 2A + 3X (3X: 2A = 1, 5 - bezvýznamné). U mužov je pomer 0, 5 2n = 2A + XY (X: 2A = 0, 5). S poklesom jeho jednotlivca ostávajú muži 3A + XY (X: 3A = 0, 33 - neplodný) - supermeni. Hodnota koeficientu medzi 1 a 0, 5 zodpovedá fenotypu jedincov prechádzajúcich pohlavím _ intersex: 3A + 2X (2X: 3A = 0, 66 - príznaky oboch pohlaví sú neplodné).
Takže podstatou teórie rovnováhy je, že nielen sexuálne chromozómy, ale aj autozómy sa zúčastňujú na určovaní pohlavia. Jedna haploidná sada autozómov hovorí o jednotlivých mužských vlastnostiach. V tomto prípade je podlaha určená pomerom počtu (bilancia) autozómov a pohlavných chromozómov.
Stanovenie pohlavia pomocou ploidy sa vyskytuje aj vo včelách. Samice sú diploidné a samce sú haploidné, pretože vyvinúť parthenogenetiku z neoplodnených vajíčok.
ROZDELENIE PODLAHY V ONTOGENÉZE
Začiatky gonád v skorých embryách (pred 5. alebo 6. týždňom) sa nelíšia v rôznych pohlavách a pozostávajú z vonkajšej vrstvy, kortexovej kôry a vnútornej vrstvy, medulky a neobsahujú bunky zárodočnej línie. Primárne bunky zárodočnej línie sa nachádzajú u ľudí v 3. týždni embryonálneho vývoja v ektoderme žĺtkového vaku. Potom pod vplyvom chemotaktických signálov migrujú do gonád. Táto migrácia je nezávislá od pohlavia. Zložky gonád sa môžu vyvinúť vo vaječníkoch alebo semenníkoch. Diferenciácia gonád sa pozoruje 8. týždeň: v 36. deň začína semenník vylučovať androgény (testosterón), ktoré určujú vývoj mužského pohlavia.
V organizmoch, ktoré majú sexuálne žľazy, sa vykonáva genetická kontrola tvorby sexuálnych charakteristík.
Rodová diferenciácia je naprogramovaná genetickou sadou pohlavných chromozómov, ktoré vznikli počas sútoku samíc a samcov. Genetický pohlavie embrya je determinované pohlavnými chromozómami XX alebo XY.
Smer vývoja pohlavia je determinovaný prítomnosťou Y chromozómu. Bežne X chromozómy obsahujú represorový gén (Tfm, gén testikulárnej feminizácie), ktorý zabraňuje vývoju mužského typu. Normálna génová alela určuje syntézu proteínového receptora pre androgény, syntetizované v obidvoch pohlaví. Vývoj fenotypu mužov závisí od génu chromozómu Y, HY-antigénu (opísaného v myši v roku 1955, génu HA). Vylučuje sa primárnymi bunkami zárodočnej línie samcov. Y 5NA 0 zodpovedá za produkciu testosterónu (mužský pohlavný hormón). Akonáhle tieto bunky vstupujú do začiatku gonád, začína diferenciácia semenníkov. Receptory HY sú prítomné na povrchu oboch typov gonadových buniek (odchýlka od vývoja pohlavia u kráv). Predpokladá sa, že samčí fenotyp je určený celým mužským chromozómom. Ale v roku 1990 bol tento gén objavený (Sex Region Y) lokalizovaný v karyotype chromozómu Y. Pri jeho absencii genotyp XY dáva ženský fenotyp.
VÝVOJ GONÁDOV
│ ┌┬┬┬ │ Gonad germ bisexuál
│ ├┼┼┼┤ │ talnaya
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
ak je genotyp XX │ │, ak je genotyp XY
(7-8 týždňov) ┴──┴──┐ ┌ ───────────────────────────────── 6
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
kortikálna │ └┴┘ │└┴┴┴┴┴┘│ medulárna vrstva
└──────────────────────────────────────────────────
(gén definuje ženský pohlavie) (gén definuje mužské pohlavie)
U ľudí, konkrétne ženské hormóny, ktoré regulujú funkcie
tieto znaky sa stávajú viditeľnými, sú produktom intersticiálnej
nás na konci 7-8 týždňov. Na 20. tkanive buniek vaječníkov alebo spermy
primárne zárodočné bunky bunky (Leydigove bunky). Tvoria
hormonálny sex (testosterón,
gonády a diferencované na estradiol). 10-12 týždňov pre-
oogonyev do konca druhého mesiaca. vnútorných pohlavných organizmov
Do konca tretieho mesiaca do hĺbky, v 12. týždni, s určitými
gonádové embryá sa zistia na úrovni androgénov v embryu
oocyty (profáza MI). Začína diferenciálna maskulinizácia (
ovariálne cysty do veku 7 mesiacov (stredný výstup) a dokončí sa
tiež sa taví do 9. mesiaca vo vaječníku do 20. týždňa.
existuje 200-400 tisíc oocytov II. V puberte je úroveň est
rogenov sa zvyšuje a ovplyvňuje
štruktúra kostry, ako aj
androgény (podľa
mužský a mužský typ).
Gonády určujú vývoj primárnych sexuálnych charakteristík a sekundárnych. Sexuálne žľazy vylučujú hormóny (estradioly, androgény), ktoré spolu s hormónmi endokrinných žliaz kontrolujú cesty sexuálnej diferenciácie. Hladina hormónov je zase riadená génmi.
Takže proces sexuálnej diferenciácie zahŕňa:
1) genetická kontrola;
2) regulačné funkcie hormónov.
Existuje teória pôsobenia hormónov ako regulačných faktorov na génoch. Pôsobia len na špecifických cieľových bunkách. V bunke sa produkuje špeciálny proteín - receptor, ktorý sa viaže na hormón a súčasne sa mení, a potom nadobúda vlastnosti na vyvolanie práce jedného alebo viacerých génov v chromozómoch. Keď glukokortikoidy pôsobia na bunky maternice, v nich sa mení syntéza RNA a proteínov (pozri schému).
Tvorba proteínových receptorov a hormónov je riadená génmi. V prípade porušenia kontroly sú možné anomálie, príkladom ktorého je Morrisov syndróm. Testikulárna feminizácia (Morrisov syndróm) (Morris, 1953): testosterónové receptory chýbajú u jedincov s touto chorobou. Androgény sú vylučované v normálnych množstvách. Pri Mauricovom syndróme je embryogenéza základom pre semenníky, ktoré začínajú produkovať mužské pohlavné hormóny. Takéto embryá však netvoria receptorový proteín (recesívna génová mutácia), ktorá zabezpečuje citlivosť buniek rozvíjajúcich sa orgánov na mužský pohlavný hormón. Vďaka tomu prestáva vývoj mužského typu a objaví sa ženský fenotyp. Vo výnimočných prípadoch je možné tieto chyby opraviť zavedením vhodných hormónov.
Preto pôvodná genetická bisexualita je základom pre nové vymedzenie pohlavia. Mužský plod nadobúda charakteristiky ženského pohlavia. Samčia karyotyp, samica gonáda, ženský fenotyp. Podiely tela sú ženy, sú prsné žľazy, skrátená vagina, semenníky - v oblasti veľkých pyskov, inguinálneho kanála, dutiny brucha.
RIEŠENIE PODLAHY
V dôsledku expozície hormónov alebo v dôsledku patológie receptorov cieľovej bunky sa môže objaviť redefinícia pohlavia (Mauricov syndróm, odstránenie pohlavných žliaz v kohútikoch).
V prírode mnohé faktory oslabujú pôsobenie génov, ktoré riadia vývoj pohlavia. Napríklad u človeka, v gonádach jedného jedinca, semenníky a vaječníky (semenník a vaječník vyvíjajú) rovnako rozvíjajú hermaprodizmus - fenomén intersexuality.
Na základe klinických údajov existujú tri typy intersexu:
1) skutočný hermafrodizmus: prítomnosť zárodočných buniek oboch pohlaví;
2) mužský pseudohermafroditismus: existujú iba semenníky (semenníky, semenníky), ženský fenotyp;
3) ženský pseudohermafroditismus: existujú len vaječníky, mužský fenotyp.
Táto klasifikácia sa nezhoduje s cytogenetickými základňami, pretože 46 XY u mužov existujú možnosti.
Pomer pohlavia
Primárny pohlavný pomer (v čase oplodnenia by mal byť blízko 1: 1, pretože stretnutie pohlavných chromozómov je rovnako pravdepodobné). Počas vyšetrenia na ľuďoch sa zistilo, že na 100 žien zygotov sa tvorí 140 až 160 samcov. Spermatozómy obsahujúce chromozóm Y sú ľahšie, mobilnejšie a majú veľký negatívny náboj (vajcia má pozitívny náboj). Preto spermie obsahujúce Y oplodňujú vajíčko častejšie.
Sekundárne - na 100 dievčat sa narodilo 103-105 chlapcov. Životaschopnosť ženských gamét, cudzorodých proteínov mužských embryí. Do veku 20 rokov je 100 chlapcov na 100 dievčat.
Terciárne - do veku 50 rokov je 85 mužov na 100 žien a vo veku 85 rokov je 50 mužov na 100 žien. Ženské telo je viac prispôsobené, čo môže byť vysvetlené spolu s inými dôvodmi aj mozaiky ženského tela na pohlavných chromozómoch.
M. Loyonova hypotéza o ženskej mozaiky na pohlavných chromozómoch.
V roku 1949 M. Borr a C. Bertrand zistili, že v jadre nervových buniek žien sa našlo zhluk intenzívne zafarbeného chromatínu. V jadrách buniek mužov to nerozpozná. Toto zhluk sa nazýva sex chromatín (telo Borra) a predstavuje jeden inaktivovaný X chromozóm.
Na začiatku vývoja v ženskom embryí fungujú oba X chromozómy, t.j. dvakrát toľko ako muži majú gény. To môže vysvetliť vyššiu životaschopnosť ženských zygotov.
V roku 1962 M. Lyon urobil hypotézu o inaktivácii jedného X chromozómu v ženskom tele cicavca. V ženskom embryu fungujú obidva chromozómy až do 16. dňa embryonálneho vývoja. 16. deň sa uskutočňuje inaktivácia jedného chromozómu pri vzniku sexuálneho chromatínu. Tento proces je náhodný, takže okolo 1/2 aktívnych buniek sa zachováva materský chromozóm X 5M 0 a otec je inaktivovaný. V iných je otec aktívny (X 5O 0) a matka je inaktivovaná. Neaktivuje sa. Matematické a otcovské X chromozómy obsahujú alelické, ale nie absolútne identické gény, t.j. dominantná alela je lokalizovaná v jednom chromozóme, druhá je recesívna. Vlastníctvo ďalších génov rozširuje adaptačné schopnosti tela.
Úrovne sexuálnej schémy sexuálnej diferenciácie
odlíšenie
┌─┐ ┌┴┐
gama X └┬┘ └─┘ Y
spermie vajíčok
┌───────────────────┐
genetická chromozómia XX a XY │
└───────────────────┘
┌─────────┴─────────┐
gonadálne │ nedefinované diferencované │
Gonáda embrya
│ └───────────────────┘
Antigén HY
V období ┌───────────────────────────────────────────────────────────────────────
genitálií │ │ │1 genitálií │
dozrievanie │ │ │ │ │ známky
│ │ │ │ └────────────┘
───────────────────────────────────────────────────────────────────
│ │gonyad │ │gадыnad ├────── 7-32 týždňov
Vaječníky │ testes │ hormóny andro
│ └──────────────────────────────────────────────────────────────────────────
Leydinga
│ │ puberty │ │ ├──────────────────────────
Rozlišovanie └────────────────────────────────────────────
hormonálny gypsén hypofýzový hormón, mužský a ženský nervový nerv
Hormóny: androgény (testosterón), estrogénne spôsoby │
└─ gény (progisterón) │ pohlavie │
┌──────────────┴───────────────┐ └────────────┘
fenotypické │ Sekundárne sexuálne charakteristiky
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
psychologická │ sexuálna a behaviorálna príslušnosť │
└───────────────────────────────────────────────────┘
Ženské telo je odolnejšie voči chladu, ionizujúcemu žiareniu, emočnému preťaženiu (Ženy plačú častejšie, aktívne amíny vystupujú so slzami, čo znižuje krvný tlak).
Ak by hypotéza fungovala bez obmedzení, nedošlo k žiadnym fenotypovým rozdielom medzi zdravými ženami s dvomi X chromozómami a pacientmi s X 4 0 alebo u mužov XY / XXYY. Je zrejmé, že chromozóm X nie je úplne inaktivovaný.
Regulácia pomeru pohlaví.
Ak umiestníte spermie do konštantného elektrického poľa, dôjde k oddeleniu chromozómov X a Y. Používa sa v hospodárskych zvieratách. Je možné získať 80% potomkov požadovaného pohlavia.
Povedomie o príslušnosti k mužskému alebo ženskému pohlaviu je neoddeliteľnou súčasťou mentálneho vnímania pohlavia, duševných parametrov. Transsexualizmus - psychologický hermaphroditismus., Fenomén porušenia sexuálnej identity jednotlivca. Prípady obsedantného povedomia o ľuďoch patriacich k opačným pohlavím sú známe od dávnych čias. Takže Herodotus vo svojom tajomstve "Scythian disease" opísal muža, ktorý nielen nosil dámske šaty, ale prijal znaky charakteru opačného pohlavia. Niektorí cisári mali tendenciu sa zamaskovať: napríklad Caligula. J. D "Ark.
Rozdiel medzi anatomickými a psychickými parametrami je konflikt medzi vnútorným prostredím a sociálnym. V mužskom tele žije ženská duša a naopak. Urobte chirurgickú korekciu podlahy. Ak chcete zmeniť muža na ženu, potrebujete 1 operáciu: vagína sa tvorí z kože penisu a šourka. Od ženy, aby človek ťažšie: 3-4 operácie (odstránenie mliečnej žľazy, vytvorenie člena). Títo ľudia sú odsúdení na večné trápenie: pijú hormóny, nemajú deti. Ústav ľudskej reprodukcie a plánovania rodiny v Moskve.
Sexuálna diferenciácia zahŕňa tvorbu primárnych (gonadových) a sekundárnych sexuálnych charakteristík.
Pohlavie vo väčšine zvierat je stanovené geneticky v čase oplodnenia.
X chromozóm nesie gén testikulárnej feminizácie (Tfm), normálne
ktorej alela určuje syntézu proteínového receptora pre androgény,
ktoré sú syntetizované v samčieho i mužského organizmu.
- každý chromozóm je nefunkčný Tfm - vedomý
├──────────────────────────────────────────────────────────────────
┌────┤ └┴┴┘ ├──────┐ XY - Aj gén obsahuje 5Na-0 gén zodpovedný za
│ └──────┘ │ Syntéza antigénu, ktorý určuje
│ M │ mužský = zodpovedný za výrobu
│ │ testosterónu.
┌──────────────────────────────────────────────────────────────────────────────────────────────────────────────────────────────────
Týždeň života. V 10. týždni môže byť sex op
Určiť súbor chromozómov.
└─────┘ └───────┘
Smer vývoja je určený prítomnosťou Y chromozómu. Zvyčajne XX chromozómy obsahujú represorový gén, ktorý zabraňuje vývoju mužského typu. Tento vývoj závisí od antigénu HY (ON), gén I 5HA je zodpovedný za produkciu testosterónu. Nachádza sa v niekoľkých druhoch (opísaných myšou v roku 1955, Eychwald, Silmser). Vylučuje sa primárnymi bunkami zárodočnej línie samcov.
Normálne sa u jedincov s kombináciou pohlavných chromozómov ženské gény ovládajú gény, ktoré určujú ženský pohlavie, a mužský typ dominuje mužský pohlavný orgán.
Pohlavné orgány sú tvorené z mulleriánskych a vlkovodov, ktoré pochádzajú z primárnej obličky. U žien sa Müllerovo kanáliky vyvíjajú do vajíčkovodov a maternice a atrofia vlfových trubíc. U mužov sa kanály vlka vyvíjajú do semenných kanálov a semenných vezikúl. Steroidné hormóny testosterón a dihydrotestosterón sú syntetizované vo fetálnych testes pod vplyvom materského hormónu (chorionický gonadotropín). Tieto hormóny pôsobia na biopotentné pupene.
vonkajšie a vnútorné pohlavné orgány: vlkodvody, Mullerovy kanály a močové sínusy. Normálne mužské telo sa vyvíja, ak fungujú všetky tieto prvky. V ich neprítomnosti sa vytvárajú sexuálne charakteristiky žien. Nedokončený vývoj mužského fenotypu u mužov
genotyp (pseudohermafroditizmus mužov).
Moderná reprodukčná stratégia:
- umelé oplodnenie;
- oplodnenie in vitro;
- umelý rast embrya a jeho prenos do maternice;
- náhradná matka.
Väčšina zvierat je dvojdomých organizmov. Pohlavie možno považovať za súbor funkcií a štruktúr, ktoré poskytujú spôsob reprodukcie potomkov a prenášanie dedičných informácií. Pohlavie je najčastejšie určované v čase oplodnenia, to znamená, že karyotyp zygota hrá hlavnú úlohu pri určovaní pohlavia. Karyotyp každého organizmu obsahuje chromozómy, ktoré sú rovnaké u obidvoch pohlaví - autozómy a chromozómy, v ktorých sa samica a mužský pohlavia líšia od seba - pohlavné chromozómy. U ľudí sú "samice" pohlavné chromozómy dva X chromozómy. Počas tvorby gamét, každé vajcia dostane jeden z X chromozómov. Podlaha, v ktorej sú vytvorené gaméty rovnakého typu, ktoré nesú chromozóm X, sa nazývajú homogametické. U ľudí je ženský pohlavný homogametický. "Mužské" pohlavné chromozómy u ľudí sú X chromozóm a Y chromozóm. Počas tvorby gamét sa polovica buniek spermií získa chromozóm X, druhá polovica dostáva chromozóm Y. Podlaha, ktorá má rôzne druhy gamét sa nazýva heterogametický. U mužov je mužské pohlavie heterogametické. Ak vznikne zygota, ktorá nesie dva X chromozómy, z nej sa vytvorí ženské telo, ak chromozóm X a Y chromozóm sú samce.
U zvierat je možné rozlíšiť nasledujúce štyroch typov chromozomálneho stanovenia pohlavia.
1. Female sex - gomogameten (XX), muž - heterogametické (XY) (cicavce, najmä človeku, Drosophila).
Genetická schéma stanovenia chromozomálneho pohlavia u ľudí:
Genetická schéma stanovenia chromozomálneho pohlavia v Drosophile:
2. Ženské pohlavie je homogametické (XX), mužské pohlavie je heterogametické (X0) (orthoptera).
Genetická schéma stanovenia chromozomálneho pohlavia v púšti kobylky:
3. Ženské pohlavie je heterogametické (XY), mužské pohlavie je homogametické (XX) (vtáky, plazy).
Genetická schéma stanovenia chromozomálneho pohlavia v holube:
Yandex.DirectAll reklamy
4. Ženské pohlavie je heterogametické (X0), mužské pohlavie je homogametické (XX) (niektoré druhy hmyzu).
Stanovenie pohlavia u ľudí sa stane mechanizmom XY. Súčasne heterogametický pohlavie je mužský, homogametický - ženský. Stanovenie pohlavia je rozdelené do troch etáp: chromozomálnej, gonádovej a fenotypickej.
Dve základné pravidlá určovania pohlavia u cicavcov
Klasické embryogénne štúdie stanovili dve pravidlá na určenie pohlavia u cicavcov. Prvý z nich bol formulovaný v šesťdesiatych rokoch Alfredom Jostom na základe pokusov odstrániť zárodok budúcich gonád (gonádový valec) v skorých embryách králikov: odstránenie valčekov pred vznikom gonády viedlo k vývoju všetkých embryí ako samíc, Bolo navrhnuté, aby boli vylučované gonády mužského efektorového hormónu testosterónu zodpovedného za maskulinizáciu plodov a predpovedal sa prítomnosť druhého efektora anti-Mullerovho hormónu (MIS), ktorý priamo riadil takéto anatomické transformácie. Výsledky pozorovaní boli formulované spravidla: špecializácia vyvíjajúcich sa gonád v semenníkoch alebo vaječníku určuje následnú sexuálnu diferenciáciu embrya.
Až do roku 1959 sa predpokladalo, že počet X chromozómov je najdôležitejším faktorom, ktorý kontroluje pohlavie u cicavcov. Objavenie organizmov s jediným X chromozómom, vyvíjajúcim sa ako ženy a jedinci s jedným Y chromozómom a viacerými X chromozómami, ktoré sa vyvinuli ako muži, ich donútili opustiť takéto pojmy. Druhé pravidlo pre určenie pohlavia u cicavcov bolo formulované: Y-chromozóm nesie genetickú informáciu potrebnú na určenie pohlavia u mužov.
Kombinácia vyššie uvedených dvoch pravidiel sa niekedy nazýva rastovým princípom: Chromozómový pohlavný syndróm spojený s prítomnosťou alebo neprítomnosťou chromozómu Y určuje diferenciáciu embryonálnej gonády, ktorá zase ovplyvňuje fenotypový pohlavie organizmu. Takýto mechanizmus na určenie pohlavia sa nazýva genetický. GSD) a postaviť sa proti nim na základe riadiacej úlohy environmentálnych faktorov (Eng. ESD) alebo pomer pohlavných chromozómov a autozómov (str. CSD).
Hormonálne stanovenie pohlavia
Definícia pohlavia môže byť reprezentovaná ako obušok, ktorý chromozomálny mechanizmus prenáša na nediferencované gonády, ktoré sa vyvinú do mužských alebo ženských pohlavných orgánov. Pri štúdiu úlohy pohlavných chromozómov vo vývoji gonád sa ukázalo, že prítomnosť alebo neprítomnosť chromozómu Y je pre človeka rozhodujúca. Pri absencii chromozómu Y dochádza k diferenciácii gonád do vaječníkov a vzniku ženy. V prítomnosti chromozómu Y sa vyvíja mužský systém. Je zrejmé, že chromozóm Y produkuje látku, ktorá stimuluje semenníkovú diferenciáciu. "Zdá sa, že základným plánom prírody bolo vytvoriť ženu a že pridanie Y-chromozómu produkuje variačný človek." V ďalšom štádiu relé pokračujú hormóny, ktoré určujú proces sexuálnej diferenciácie plodu a jeho anatomický vývoj. Pri narodení prvá časť programu končí. Po narodení prechádza obušok na environmentálne faktory, ktoré dokončujú formovanie pohlavia - zvyčajne, ale nie vždy, podľa genetického pohlavia. Stanovenie pohlavia je komplexný viacstupňový proces, ktorý na ľuďoch závisí, okrem biologického, od psychosociálnych faktorov. To môže viesť k transsexualite, heterosexuálnemu, bisexuálnemu alebo homosexuálnemu správaniu a životnému štýlu.
Fyziologický základ stanovenia gonádového pohlavia
Fyziologickým základom mechanizmu stanovenia pohlavia je bisexualita cicavcov plodných gonád. V takom progonadah súčasne predstavovať Müllerian potrubia a Wolff kanál - začiatky genitálneho traktu u mužov a žien, v danom poradí. Primárne stanovenie pohlavia začína výskytom špecializovaných bunkových línií, Sertoliho bunky. V druhom z nich sa syntetizuje anti-Müller hormón, predpovedaný Zhostom, ktorý je zodpovedný za priamu alebo nepriamu inhibíciu vývoja Müllerovho kanála, klíčika budúcich vajíčkovodov a maternice.
Genetický mechanizmus určovania pohlavia
Ľudský Y chromozóm, čo naznačuje umiestnenie génu SRY
V roku 1987 David Page a jeho kolegovia vyšetrovali XX muža, ktorý zdedil špecifický fragment dvojice Y chromozómov 280 tisíc nukleotidov a XY žena s deléciou, ktorá zachytáva túto oblasť ako výsledok výmeny chromozómov. Tento fragment je prítomný v chromozóme Y všetkých skutočných zvierat z eutherie a nachádza sa vo vzdialenosti 100 tisíc párov báz od hranice pseudoautozomálnej oblasti génu ZFY s dĺžkou 140 tisíc párov báz.
ZFY homológ - gén ZFX sa nachádza na chromozóme X a ZFX nie je inaktivovaný. Obidva faktory ZFX a ZFY kódujú transkripčné faktory obsahujúce motívy zinkových prstov s aktivitou viažucou DNA. Ďalšia podrobná analýza špecifických sekvencií Y chromozómov u jedincov s inverznou sexuou obmedzila vyhľadávanie na oblasť o veľkosti 35 kbp a viedla k objaveniu génu považovaného za skutočný ekvivalent klasickej angličtiny. Faktor určujúci Testis, Tento gén sa nazýva SRY (in. Stanovenie pohlavia Gén regiónu Y).
SRY ktorý sa nachádza v oblasti stanovenia pohlavia a obsahuje konzervatívnu doménu (HMG-box), kódujúcu proteín s 80 aminokyselinovými zvyškami. Aktivita génu SRY je zaznamenaná pred začiatkom obdobia diferenciácie priebehu do semenníka - 10-12 dňa myšieho embryonálneho vývinu a prinajmenšom v tomto štádiu nezávisí od prítomnosti zárodočných buniek. Špecifické bodové mutácie alebo delécie v HMG krabici tohto génu u žien XY vedú k sexuálnej inverzii. Prenos 14 kbp DNA fragmentu obsahujúceho tento gén so susediacimi oblasťami do oplodneného vajíčka homogametického jedinca pomocou mikroinjekcie mal za následok vyzeranie samca s karyotypom XX.
Génové funkcie SRY
Doména kódovaná HMG krabicou SRY génu sa špecificky viaže na DNA a spôsobuje jej ohnutie. Ohýbanie DNA spôsobené SRY proteínom alebo jeho homológmi obsahujúcimi doménu HMG sa môže mechanicky preniesť na značnú vzdialenosť a hrať dôležitú úlohu pri regulácii transkripcie, replikácie a rekombinácie. Oblasť DNA, v ktorej je SRY lokalizovaná, obsahuje dva gény kódujúce kľúčové enzýmy zahrnuté v primárnej diferenciácii gonád mužského typu: P450 aromatázový gén, ktorý riadi konverziu testosterónu na estradiol a inhibítor vývoja kmeňa Miller, čo spôsobuje ich opačný vývoj a podporuje diferenciáciu semenníkov ,
Génový produkt SRY sa tiež zúčastňuje procesov sexuálnej diferenciácie v úzkej interakcii s iným génom nazývaným gén Z, ktorého funkcia normálne spočíva v inhibícii špecifických mužských génov. V prípade normálneho samec genotypu 46XY SRY gén kóduje proteín, depresívne gén Z, a špecifické samčie gény sa aktivujú. V prípade normálneho ženského genotypu 46XX, v ktorom SRY chýba, je aktivovaný gén Z a inhibuje špecifický mužský gén, ktorý vytvára podmienky pre vývoj ženského typu.
Chromozomálna teória stanovenia pohlavia
Zvážte chromozómové stanovenie pohlavia. Je známe, že v dvojdomých organizmoch (zvieratá a dvojdomé rastliny) je pomer pohlaví obyčajne 1: 1, to znamená, že samci a samice sú rovnako bežné. Tento pomer sa zhoduje s rozdelením na analyzovanom kríži, keď jedna z krížených foriem je heterozygotná (Aa), a druhá je homozygotná pre recesívne alely (AA). V potomstve je v tomto prípade rozdelenie vo vzťahu k 1 Aa: laa. Ak je pohlavie zdieľané podľa toho istého princípu, potom by bolo logické predpokladať, že jeden pohlavie by malo byť homozygotné a iné heterozygotné. Rozdelenie podľa pohlavia v každej generácii by sa malo rovnať 1,1, čo je skutočne pozorované.
Evolučná teória rodov Navrhol to V. Geodakyan v roku 1965. Teória vysvetľuje z jednotného hľadiska veľa javov súvisiacich s pohlavím: sexuálny dimorfizmus je normálny a patológia je pohlavný pomer, diferenciálna úmrtnosť a miera sexuálnej odpovede, úloha pohlavných chromozómov a pohlavných hormónov, asymetria mozgu a rúk, vzájomné účinky a psychologické a sociálne rozdiely medzi pohlaviami.
Teória je založená na princípe konjugovaných subsystémov, ktoré sa vyvíjajú asynchrónne. Muž je prevádzkové subsystém populácie, konzervatívny subsystém. Nové informácie o životnom prostredí padá prvý mužského pohlavia, a to iba po mnoho generácií odovzdaný k žene, takže vývoj mužských evolúcie predchádza ženy. Tento časový posun (dva fázy vývoj znaku) vytvára dve formy znaku (mužského a ženského) - pohlavný dimorfizmus v populácii. Evolučná "vzdialenosť" medzi subsystémami je potrebná na vyhľadávanie a kontrolu inovácií.
 Ako boli vynájdené listy a abeceda - história vývoja listu
Ako boli vynájdené listy a abeceda - história vývoja listu Zriedkavé prírodné javy, o ktorých vie len málo ľudí
Zriedkavé prírodné javy, o ktorých vie len málo ľudí Aký je názov osoby, ktorá zmenila pohlavie
Aký je názov osoby, ktorá zmenila pohlavie