Je li ženski spol uvijek određen kombinacijom xx. Podna biologija
Genetika poda
Spol karakterizira kompleks obilježja određenih genima smještenim u kromosomima. U stanicama ljudskog tijela, kromosomi su upareni diploidni skupovi. Kod vrsta s dvodomnim osobama, kromosomski kompleks mužjaka i ženki nije isti i razlikuje se jednim parom kromosoma (spolni kromosomi). Identični kromosomi ovog para nazvani su X (X) -kromosomom, nespareni, odsutni iz drugog kromosoma spola - Y (igra); ostatak za koji ne postoje razlike su autosomi (A).
Ženske stanice sadrže dva identična spolna kromosoma, označena XX, u muškaraca su predstavljena s dva nesparena kromosoma X i Y. Dakle, skup kromosoma muškarca i žene razlikuje se samo u jednom kromosomu: kromosomski skup žene sadrži 44 autosoma + XX, muškarci - 44 autosome + XY.
Tijekom podjele i sazrijevanja zametnih stanica kod ljudi nastaju gamete s haploidnim brojem kromosoma: jaja u pravilu sadrže 22 + X kromosoma. Tako se kod žena formira samo jedna vrsta gameta (gamete s X kromosomom). Kod muškaraca, gamete sadrže 22 + X ili 22 + Y kromosoma, te nastaju dvije vrste gameta (gameta s X kromosomom i gameta s Y kromosomom). Ako tijekom oplodnje spermatozoid s X kromosomom uđe u jajnu stanicu, formira se ženska klica, a Y kromosom je muški.
Stoga, određivanje spola osobe ovisi o prisutnosti u muškim zametnim stanicama - spermi, gnojidbi jaja, X- ili Y-kromosoma.
Postoje četiri glavne vrste određivanja kromosomskog spola:
1. Muški spol je heterogametan; 50% gameta nosi X-, 50% -U - kromosoma, na primjer, čovjeka, sisavaca, diptera, kukaca, kukaca (Slide 4).
2. Muški spol je heterogametan; 50% gameta su X-, 50% nemaju spolni kromosom, na primjer skakavci, klokani (Slide 7).
3. Ženski spol je heterogametan; 50% gameta nosi X-, 50% gameta -Y- kromosoma, na primjer, ptice, gmazovi, repovi vodozemci, svilena buba (Slide 7).
4. Ženski spol je heterogametan; 50% gameta nosi X-, 50% nema spolni kromosom, primjerice krticu.
Nasljeđivanje osobina čiji su geni lokalizirani u spolnim kromosomima naziva se nasljeđivanjem, spojen s podom.
26. Genotip kao integralni sustav. Interakcija gena, višestruko djelovanje gena.
Genotip sličan holistički sustav
Svojstva gena. Na temelju u potrazi s primjerima nasljeđivanja osobina u mono- i di-hibridnim križevima, može se steći dojam da je genotip organizma sastavljen od zbroja odvojenih, neovisno djelujućih gena, od kojih svaki određuje razvoj vlastite osobine ili svojstva. Takva ideja izravne i nedvosmislene povezanosti gena s osobinom najčešće ne odgovara stvarnosti. Zapravo, postoji veliki broj znakova i svojstava živih organizama, koji su određeni s dva ili više parova gena, i obrnuto, jedan gen često kontrolira mnoge znakove. Osim toga, učinak gena može se promijeniti zbog blizine drugih gena i uvjeta okoline. Dakle, u ontogenezi ne djeluju pojedinačni geni, već cijeli genotip kao integralni sustav sa složenim vezama i interakcijama između njegovih komponenti. Ovaj sustav je dinamičan: pojava novih alela ili gena kao posljedica mutacija, formiranje novih kromosoma, pa čak i novih genoma dovodi do zamjetne promjene genotipa tijekom vremena.
Priroda manifestacije djelovanja gena u sastavu genotipa kao sustava može varirati u različitim situacijama i pod utjecaj različitih čimbenika. To se može lako vidjeti ako uzmemo u obzir svojstva gena i karakteristike njihove manifestacije u znakovima:
Gen je diskretan u svom djelovanju, tj. Izoliran je u svojoj aktivnosti od drugih gena.
Gen je specifičan u svojoj manifestaciji, tj. Odgovoran je za strogo određenu karakteristiku ili svojstvo organizma.
Gen može djelovati postupno, tj. Povećati stupanj manifestacije svojstva s povećanjem broja dominantnih alela (doza gena).
Jedan gen može utjecati na razvoj različitih osobina - to je višestruki ili pleiotropni učinak gena.
Različiti geni mogu imati isti učinak na razvoj iste osobine (često kvantitativna svojstva) - to su višestruki geni ili poligeni.
Gen može stupiti u interakciju s drugim genima, što dovodi do pojave novih znakova. Takva interakcija provodi se neizravno - kroz produkte njihovih reakcija sintetiziranih pod njihovom kontrolom.
Djelovanje gena može se modificirati promjenom njegovog položaja na kromosomu (efekt pozicije) ili utjecajem različitih čimbenika okoline.
Interakcije alelnih gena. Fenomen kada je nekoliko gena (alela) odgovorni za jednu osobinu naziva se interakcija gena. Ako su to aleli istog gena, onda se takve interakcije nazivaju alela, i u slučaju alela različitih gena - non-alelnu.
Razlikuju se sljedeći glavni tipovi alelnih interakcija: dominacija, nepotpuna dominacija, naddominacija i suodnosnost.
dominacija - tip interakcije dvaju alela jednog gena, kada jedan od njih potpuno eliminira manifestaciju drugog. Takav fenomen moguć je pod sljedećim uvjetima: 1) dominantni alel u heterozigotnom stanju osigurava sintezu produkata dovoljnih za ispoljavanje osobine iste kvalitete kao u stanju dominantnog homozigota u roditeljskom obliku; 2) recesivni alel je potpuno neaktivan, ili produkti njegove aktivnosti ne djeluju s proizvodima aktivnosti dominantnog alela.
Primjeri ove interakcije alelnih gena su dominacija ljubičaste boja cvijeće graška preko bijele, glatke strukture sjemena nad naboranom, tamnom kosom iznad svjetla, smeđe oči u plavoj boji ljudi, itd.
Nepotpuna dominacija ili posredni karakter nasljeđivanja uočeno u slučaju kada se fenotip hibrida (heterozigota) razlikuje od fenotipa obaju roditeljskih homozigota, tj., ekspresija osobine je intermedijarna, s više ili manje odstupanja prema jednom ili drugom roditelju. Mehanizam ove pojave je u tome što je recesivni alel neaktivan, a stupanj aktivnosti dominantnog alela je nedovoljan da osigura željenu razinu manifestacije dominantne osobine.
Primjer nepotpune dominacije je nasljeđivanje. boja cvijeće u biljkama noćne ljepote (sl. 3.5). Kao što se može vidjeti iz dijagrama, homozigotne biljke imaju ili crvenu (AA) bilo bijelo (Aa) cvjetovi i heterozigotni (Aa) - ružičasta. Kada prelazimo biljke s crvenim cvjetovima i biljkama s bijelim cvjetovima u F 1, sve biljke imaju ružičaste cvjetove, to jest, postoji posredni karakter nasljeđivanja. Kod križanja hibrida s ružičasto cvijeće F 2 postoji podudarnost cijepanja fenotipom i genotipom, budući da je dominantni homozigot (AA) različiti od heterozigota (Aa). Dakle, u ovom primjeru s noćnim ljepotama, cijepanjem F 2 Boja cvijeća je obično sljedeća - 1 crvena (AA): 2 roze (Aa): 1 bijelo (Aa).
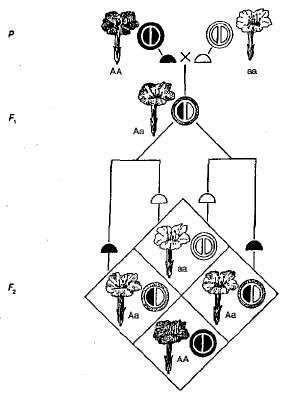
Sl. 3.5. Nasljeđivanje boje cvijeća u slučaju nepotpune dominacije noćne ljepote.
Nepotpuna dominacija postala je raširena. Uočava se u nasljeđivanju kovrčave kose kod ljudi, boji goveda, boji perja kod pilića i mnogim drugim morfološkim i fiziološkim karakteristikama biljaka, životinja i ljudi.
ogledima - jača manifestacija svojstva u heterozigotnom pojedincu (Aa), nego bilo koji homozigot (AA i aa). Pretpostavlja se da je ovaj fenomen u osnovi heterozisa (vidi § 3.7).
Kodaminirvanie- sudjelovanje oba alela u određivanju svojstva u heterozigotnom pojedincu. Upečatljiv i dobro proučen primjer suživota je nasljeđivanje krvne skupine IV kod ljudi (skupina AB).
Crvene krvne stanice u ovoj skupini imaju dvije vrste antigena: antigen (određeno genomom / dostupnim u jednom od kromosoma) i antigenom (određen genomom / a, lokaliziran na drugom homolognom kromosomu). Samo u ovom slučaju oba alela - 1 (U homozigotno stanje kontrolira krvnu skupinu II, skupinu A) i ja B (u homozigotnom stanju kontrolira III krvnu skupinu, skupina B). aleli 1 i ja B rad u heterozigotu kao da su međusobno neovisni.
Primjer nasljeđivanja grupe krv ilustrira i manifestira višestruki alelizam: Gen / može biti predstavljen s tri različita alela, a postoje geni koji imaju na desetine alela. Imenovani su svi aleli istog gena niz višestrukih alela, od kojih svaki diploidni organizam može imati bilo koja dva alela (i samo). Između tih alela moguće su sve navedene varijante alelnih interakcija.
Fenomen višestrukog alelizma je čest u prirodi. Postoji opsežna serija višestrukih alela koji određuju vrstu kompatibilnosti tijekom gnojidbe gljivicama, oprašivanje u sjemenskim biljkama, određivanje boje životinjske dlake itd.
Interakcije nealelnih gena Interakcije gena koje nisu alele opisane su u mnogim biljkama i životinjama. Oni dovode do pojave u potomstvu digerozigota neuobičajenog cijepanja prema fenotipu: 9: 3: 4; 9: 6: 1; 13: 3; 12: 3: 1; 15: 1, tj. modifikacije opće Mendelijanove formule 9: 3: 3: 1. Postoje slučajevi interakcije dvaju, tri i više nealelnih gena. Među njima su sljedeći glavni tipovi: komplementarnost, epistaza i polimeri.
komplementarni ili dodatne, Ta se interakcija naziva nonallelnim dominantnim genima, zbog čega se pojavljuje osobina koja nije prisutna kod oba roditelja. Na primjer, kada se dvije vrste slatkog graška prekriže s bijelim cvjetovima, potomci se pojavljuju s ljubičastim cvjetovima. Ako označite genotip jedne sorte AAbb, i drugo aABB,
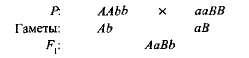
Hibrid prve generacije s dva dominantna gena (A i u) Dobio je biokemijsku osnovu za proizvodnju antocijanin purpurnog pigmenta, dok je jedan po jedan gen i, Niti jedan gen B nije osigurao sintezu tog pigmenta. Sinteza antocijana je složeni lanac sekvencijalnih biokemijskih reakcija koje kontrolira nekoliko nealelnih gena i samo ako postoje najmanje dva dominantna gena (A-B) razvija se ljubičasta boja. U drugim slučajevima (aaV- i A-bb) cvjetovi biljke su bijeli (znak "-" u formuli genotipa znači da ovo mjesto može zauzeti i dominantni i recesivni alel).
Od samoprašivanja biljaka slatkog graška F 1 u F 2 podjela na grimizne i bijelo obojane oblike uočena je u omjeru blizu 9: 7. U njemu su pronađeni ljubičasti cvjetovi 9/1 6 biljaka, bijelo - u 7/16. Pennetova rešetka jasno pokazuje uzrok ove pojave (sl. 3.6).
Epistaza - ovo je vrsta interakcije gena u kojoj aleli jednog gena potiskuju manifestaciju alelnog para drugog gena. geni inhibiraju djelovanje drugih gena epistatski inhibitori ili prigušivači. Potisnut gen se naziva hipostatska.
Prema promjeni broja i omjera fenotipa i cikličkih klasa tijekom cijepanja dihibrida F 2 razmotriti nekoliko vrsta epistatskih interakcija: dominantna epistaza (A\u003e B ili B\u003e A) s cijepanjem 12: 3: 1; recesivna epistaza (a\u003e B ili b \u003e A), koja je izražena u cijepanju 9: 3: 4, itd.
polymerism što se očituje u činjenici da je jedan znak formiran pod utjecaj nekoliko gena s istom fenotipskom ekspresijom. Ti se geni nazivaju polimer. U ovom slučaju usvojen je princip nedvosmislenog djelovanja gena na razvoj svojstva. Primjerice, kada prelaze biljke s pastirskim vrećicama s trokutastim i ovalnim plodovima (mahuna) u F 1, formiraju se biljke s plodovima trokutastog oblika. S njihovim samooprašivanjem F 2 Uočava se cijepanje na biljke s trokutastim i ovalnim mahunama u omjeru 15: 1. To se objašnjava činjenicom da postoje dva gena koji djeluju jedinstveno. U tim se slučajevima označavaju identično. 1 i 2 .
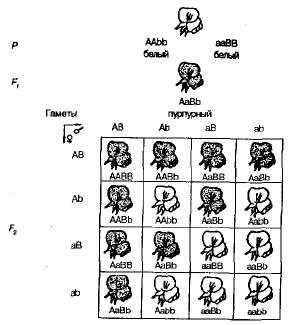
Sl. 3.6 , Nasljeđivanje boje cvijeća u slatkom grašku
Zatim svi genotipovi (A 1 ,- 2 ,-, 1 a 2 i 2 A 1 1 2 -) imat će isti fenotip - trokutaste mahune i samo biljke i 1 i 1 i 2 2 će biti drugačiji - formirati ovalne mahune. To je slučaj ne-kumulativni polimeri.
Polimerni geni mogu djelovati prema tipu kumulativni polimeri. Što je više sličnih gena u genotipu organizma, to je jača manifestacija te osobine, tj. S povećanjem doze gena (A 1 2 3 itd.) njegovo djelovanje je sažeto ili kumulirano. Primjerice, intenzitet boje endosperma zrna pšenice proporcionalan je broju dominantnih alela različitih gena u trihibridnom križanju. Najviše obojenih zrnaca su 1 1 2 2 3 , A 3 i zrno a 1 i 1 i 2 2 i 3 i 3 nije imao pigment.
Prema vrsti kumulativnog polimera, mnogi se znakovi nasljeđuju: mliječnost, proizvodnja jaja, težina i drugi znakovi domaćih životinja; mnoge važne parametre fizičke snage, zdravlja i mentalnih sposobnosti osobe; duljina zrna uha; sadržaj šećera u korijenu ili lipidima šećerne repe suncokretovo sjeme i tako dalje
Prema tome, brojna zapažanja upućuju na to da je manifestacija većine znakova posljedica utjecaja kompleksa interakcijskih gena i uvjeta okoliša na formiranje svake specifične osobine.
Interakcija gena
Odnos između gena i osobina je prilično kompliciran. U organizmu, ne uvijek jedan gen određuje samo jednu osobinu i, obrnuto, jednu osobinu određuje samo jedan gen. Češće, jedan gen može pridonijeti ispoljavanju nekoliko znakova odjednom, i obrnuto. Genotip organizma ne može se smatrati jednostavnim zbrojem neovisnih gena, od kojih svaki funkcionira izvan dodira s drugima. Fenotipske manifestacije osobina rezultat su interakcije mnogih gena.
Višestruko djelovanje gena (pleiotropija) - procesi utjecaja jednog gena na nastanak nekoliko znakova.
Na primjer, kod ljudi, gen koji određuje crvenu boju kose uzrokuje svjetliju kožu i pjege.
Ponekad geni koji određuju morfološke značajke utječu na fiziološke funkcije, smanjuju vitalnost i plodnost ili su smrtonosni. Dakle, gen koji uzrokuje plavu boju u minki smanjuje njegovu plodnost. Detaljan je dominantni gen sive boje u arakulskim ovcama u homozigotnom stanju, budući da takva janjad ima nerazvijen želudac i umiru kada prelaze u travu.
Komplementarna interakcija gena. Nekoliko gena može utjecati na razvoj jedne osobine. Interakcija nekoliko nealelnih gena, koja dovodi do razvoja jedne osobine, naziva se komplementarna. Primjerice, kokoši imaju četiri oblika grba, manifestacija bilo koje od njih povezana je s interakcijom dva para nealelnih gena. Ružičasti greben je posljedica djelovanja dominantnog gena jednog alela, grebena u obliku graška - dominantnog gena drugog alela. Kod hibrida, u prisutnosti dva dominantna nealelna gena, formira se orahov nalik, au odsustvu svih dominantnih gena, tj. u recesivnim homozigotima za dva nonallelna gena nastaje jednostavan češalj.
Rezultat interakcije gena je bojanje vune kod pasa, miševa, konja, oblika bundeve, boje cvijeća slatkog graška.
Polimer je interakcija nealelnih gena kada stupanj razvoja osobine ovisi o ukupnom broju dominantnih gena. Prema tom principu, naslijeđuje se bojenje zrna zobi i pšenice i boja kože osobe. Na primjer, u crncima postoje 4 dominantna gena u dva para nealelnih gena, a nitko u ljudima bijele kože, svi geni su recesivni. Kombinacije različitih količina dominantnih i recesivni geni dovesti do stvaranja mulata s različitim intenzitetom boje kože: od tamne do svjetle.
Postoje dvije glavne skupine interakcije gena: interakcija između alelnih gena i interakcija između ne-alelnih gena. Međutim, treba shvatiti da to nije fizička interakcija samih gena, nego interakcija primarnih i sekundarnih produkata koji će odrediti jednu ili drugu osobinu. U citoplazmi se javlja interakcija između proteina - enzima čija sinteza je određena genima ili između tvari koje nastaju pod utjecajem tih enzima.
Moguće su sljedeće vrste interakcija:
1) za formiranje specifične osobine potrebna je interakcija dvaju enzima, čija se sinteza određuje s dva ne-alelna gena;
2) enzim koji je sintetiziran uz sudjelovanje jednog gena potpuno suzbija ili inaktivira djelovanje enzima, koji je nastao drugim nealelnim genom;
3) dva enzima, čije je formiranje kontrolirano s dva ne-alelna gena koji utječu na jednu osobinu ili jedan proces tako da njihovo zajedničko djelovanje dovodi do nastanka i poboljšanja manifestacije osobine.
Interakcija alelnih gena
Geni koji zauzimaju identične (homologne) lokuse u homolognim kromosomima nazivaju se alelni. Svaki organizam ima dva alelna gena.
Poznati su takvi oblici interakcija između alelnih gena: potpuna dominacija, nepotpuna dominacija, kodominantnost i naddominancija.
Glavni oblik interakcije je potpuna dominacija, koju je prvi opisao G. Mendel. Njegova suština leži u činjenici da u heterozigotnom organizmu ispoljavanje jednog od alela dominira nad manifestacijom drugog. Uz potpunu dominaciju cijepanja genotipom 1: 2: 1 ne podudara se s cijepanjem fenotipom - 3: 1. U medicinskoj praksi, od dvije tisuće monogenih nasljednih bolesti, gotovo polovica ima dominantnu manifestaciju patoloških gena u odnosu na normalne. U heterozigotima se patološki alel očituje u većini slučajeva znakovima bolesti (dominantni fenotip).
Nepotpuna dominacija je oblik interakcije u kojem dominantni gen (A) u heterozigotnom organizmu (Aa) ne potiskuje u potpunosti recesivni gen (a), zbog čega se pojavljuje međuprostor između roditeljskih znakova. Ovdje je podjela genotipa i fenotipa ista i iznosi 1: 2: 1
Kada je kodominantan u heterozigotnim organizmima, svaki od alelnih gena uzrokuje stvaranje produkta koji ovisi o njemu, odnosno ispada da su proizvodi oba alela. Klasičan primjer takve manifestacije je sustav krvnih grupa, posebno sustav ABO, kada ljudski eritrociti nose antigene na površini koji su pod kontrolom oba alela. Ovaj oblik manifestacije naziva se ko-dominacija.
Superdominacija - kada je dominantni gen izraženiji u heterozigotnom stanju nego u homozigotnom stanju. Tako u Drosophili, s genotipom AA, normalni očekivani životni vijek; Aa - produženi trivatistički život; aa - smrtonosna.
Više navoda
Svaki organizam ima samo dva alelna gena. Međutim, često u prirodi, broj alela može biti više od dva, ako neki lokus može biti u različitim stanjima. U takvim slučajevima, kažu se višestruki aleli ili višestruki aleomorfizmi.
Višestruki aleli označeni su istim slovom s različitim indeksima, primjerice: A, A1, A3 ... Alelni geni nalaze se u istim područjima homolognih kromosoma. Budući da u kariotipu uvijek postoje dva homologna kromosoma, čak i kod višestrukih alela, svaki organizam može imati samo dva identična ili različita alela u isto vrijeme. Samo jedna od njih ulazi u reproduktivnu stanicu (zajedno s razlikom u homolognim kromosomima). Za višestruke alele, karakteristični učinak svih alela na istu osobinu. Razlika između njih leži samo u stupnju razvoja osobine.
Druga značajka je da somatske stanice ili stanice diploidnih organizama sadrže najviše dva alela od nekoliko, budući da se nalaze u istom lokusu kromosoma.
Još jedna značajka je svojstvena višestrukim alelima. Po prirodi dominacije, alelomorfni znakovi su smješteni u uzastopnom redu: češće normalna, nepromijenjena osobina dominira nad ostalima, drugi gen serije je recesivan u odnosu na prvi, ali dominira sljedećim, itd. Primjer manifestacije višestrukih alela u osobi su krvne skupine ABO sustava.
Višestruki alegizam ima važnu biološku i praktičnu važnost, jer povećava kombinacijsku varijabilnost, osobito genotipsku.
Interakcija nealelnih gena
Postoje mnogi slučajevi u kojima su svojstva ili svojstva određena s dva ili više nerazdvojnih gena koji međusobno djeluju. Iako je ovdje interakcija uvjetovana, jer ne djeluju geni, nego proizvodi koje kontroliraju. U ovom slučaju postoji odstupanje od Mendelovih uzoraka cijepanja.
Postoje četiri glavne vrste interakcije gena: komplementarnost, epistaza, polimeri i modificirajuće djelovanje (pleiotropija).
Komplementarnost je takva vrsta interakcije nealelnih gena, kada jedan dominantni gen dopunjuje djelovanje drugog neleelno dominantnog gena, a zajedno definiraju novu osobinu koja je odsutna kod roditelja. Štoviše, odgovarajuća osobina se razvija samo u prisutnosti oba ne-alelna gena. Na primjer, sumpor je boja vune u miševa kojima upravljaju dva gena (A i B). Gene A određuje sintezu pigmenta, međutim, i homozigoti (AA) i heterozigoti (Aa) su albino. Drugi gen B daje nakupine pigmenta uglavnom na bazi i na vrhovima kose. Križanje diheterozigota (AaBb x AaBb) dovodi do cijepanja hibrida u omjeru 9: 3: 4. Numerički omjeri za komplementarne interakcije mogu biti 9: 7; 9: 6: 1 (modifikacija Mendeljejevog cijepanja).
Primjer komplementarne interakcije gena u ljudi može biti sinteza zaštitnog proteina, interferona. Njegovo formiranje u tijelu povezano je s komplementarnom interakcijom dvaju nealelnih gena smještenih u različitim kromosomima.
Epistaza je interakcija nealelnih gena u kojima jedan gen potiskuje djelovanje drugog nealelnog gena. I dominantni i recesivni geni mogu uzrokovati depresiju (A\u003e B, A\u003e B, B\u003e A, B\u003e A), a ovisno o tome, epistaza je dominantna i recesivna. Prevladavajući gen naziva se inhibitor ili supresor. Geni inhibitora općenito ne određuju razvoj određene osobine, već samo inhibiraju djelovanje drugog gena.
Gen čiji je učinak potisnut naziva se hipostatičan. S epistatičnom interakcijom gena, fenotipsko cijepanje u F2 je 13: 3; 12: 3: 1 ili 9: 3: 4, itd. Boja plodova bundeve, boja konja određena je ovom vrstom interakcije.
Paul - to je skup morfoloških, fizioloških, biokemijskih, bihevioralnih i drugih znakova tijela koji uzrokuju reprodukciju.
Znakovi po kojima se razlikuju različiti spolovi dijele se na primarne i sekundarne. Primarni organi su organi koji osiguravaju stvaranje gameta i oplodnju (gonade, genitalni trakt, organe). To su vanjski i unutarnji genitalni organi, koji se polažu u embriogenezu. Sekundarno - ne sudjelujte u spolnoj reprodukciji. Razvijaju se pod utjecajem spolnih hormona i pojavljuju se u pubertetu (kod ljudi za 12-15 godina). To su obilježja razvoja mišićno-koštanog sustava, potkožnog masnog tkiva, kose, ton glasa, osobine ponašanja; u pticama, pjevanje, perje itd.
Znakovi osoba povezanih sa spolom mogu se podijeliti u 3 kategorije:
1) ograničeno na pod
2) podno kontrolirano
3) spojen s podom.
Razvoj prvog je posljedica gena koji se nalaze u autozomima oba spola, ali se manifestiraju samo u jednom spolu. Tako su geni za polaganje jaja prisutni u pilićima i pijetlovima, ali se pojavljuju samo kod pilića. Slično se ponašaju i geni masnoće i mlijeka u goveda. Ovaj fenomen je uočen u vezi s izlaganjem odgovarajućim spolnim hormonima.
Primjer drugog je manifestacija rogova kod muških krava, komolost - u ženki. Kod ljudi: ćelavost kod muškaraca, giht - 80% u muškaraca i 12% u žena.
Znakovi, razvoj kojih kontroliraju geni spolnih kromosoma, nazivaju se spolno povezani. Takvih znakova ima oko 200. Kod X kromosoma postoji povezana sljepoća za boje, hemofilija; Y-kromosom - hipertrihoza, ihtioza.
VRSTE ODREĐIVANJA TLA.
1. Progamous - prije oplodnje. Odnos spolnih kromosoma u ovom slučaju ne igra ulogu, jer diploidne oocite. (Neki crvi, rotiferi - ženke se razvijaju iz velikih oocita, mužjaka iz malih jajnih stanica).
2. Singamsko - genetsko određivanje spola tijekom oplodnje, koje ovisi o prirodi kombinacije spolnih kromosoma ili omjeru spolnih kromosoma i autosoma.
3. Epigamic - pod utjecajem vanjske okoline (bonelia crv).
KROMOSOMALNA DEFINICIJA KATU
┌─────────────────────────┬───────────────────┬──────────────────┐
│ Kromosomski tipovi oty genotipovi │ tipovi gameta
├ određivanje spola ├─────────┬─────────┼────────┬─────────┤
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
Hete muški heterogame │
Inse Kukci za rode │ │ │ │ │
Bug (kukci Protenor, kukci, │ │ │ │)
Ukipauki, skakavci) │ HO │ HH, H, O │ H │
Oph Drosophilus │ XY │ XX, X, Y X │
Br Kičmenjaci br │ │ │
Mam (sisavci, čovjek) Y XY │ XX, X, Y │ X │
│ │ │ │ │ │
Hete ženski heterogamma │
S Ptice. ribe, leptiri, │ │ │ │
│ kop │ │ │ │
. Vodeni. │ XX Y XY │ X, X, Y │
AndCijena i ostali nemoćni │ │ │ │
Noć │ XX O XO │ X │ X, O
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
Sinkamički tip definicije je definicija kromosoma genima s genetskom kontrolom. Odgovoran za spol kromosoma koji se zove seks. Normalni muški gamet nosi ili X ili Y kromosom, a sva jaja - X kromosom. U slučaju normalnog kromosomskog odstupanja tijekom mejoze nastaju normalna jajašca i sperma s uobičajenim setom kromosoma X i Y. Pol zigota određuje se omjerom gameta XX i XY (homogametni i heterogametni).
Corrensova kromosomska teorija spola (1907.) je da je spol određen kombinacijom spolnih kromosoma tijekom oplodnje. Razlikuju se sljedeće vrste određivanja kromosomskog spola: XY, XO, ZW, ZO.
U slučaju narušavanja tijeka mitoze ili mejoze, mogu se formirati ginandomorfi. Sadržaj spolnih kromosoma u različitim stanicama takvih pojedinaca može biti različit (mozaik). U Drosophili lete: XX i XO, u čovjeku XX i XY, u vezi s kojima različite boli u tijelu mogu imati odgovarajuće znakove spola. Mogu postojati i drugi slučajevi mozaika: XX / XXX, XY / XXX; XO / XXY i drugi
Ako se spolni kromosomi ne rastave, može postojati 12 mogućih kombinacija spolnih kromosoma u ljudskim zigotama, što je uzrok kromosomskih aberacija kod ljudi.
│ X │ XX │ O
─────┼────────┼─────────┼────────
X │ XX │ XXX O XO
Y Y XY │ XXY. YO
XY Y XXY Y XXXY YO XYO
O O XO │ │
U slučaju nedisjunkcije spolnih kromosoma tijekom mejoze, gamete XX i O nastaju u ženki. i XY i O kod muškaraca. Svojim sudjelovanjem u oplodnji, zigoti se formiraju s neobičnom kombinacijom spolnih kromosoma. Kod ljudi se takve anomalije javljaju 1 u 600-700 novorođenčadi. Zygote Yo umire u ranoj fazi; pojedinci XXX, XXY, XO su održivi i njihov spol ovisi o prisutnosti ili odsutnosti "Y" kromosoma, koji, s bilo kojim brojem X kromosoma, kontrolira stvaranje znakova muškog spola, razvoj i stimulira formiranje testisa. Višak X kromosoma uzrokuje ustavne anomalije i nedostatke inteligencije. No, u prirodi postoje pojedinci u kojima je „Y“ kromosom genetski inertan i nema poseban utjecaj na određivanje spola.
Nađeni su uzorci vrste Drosophila tipa CW, koji su bili mužjaci, ali neplodni (1916, C. Mostovi), a XXY pojedinci su normalne plodne žene.
Teorija ravnoteže spola (Bridges, 1922). Proučavala je omjer genitalija i autosoma
Kod normalnih ženki sa skupom kromosoma 2n, omjer autosoma i X kromosoma je 1: 2n = 2A + 2X (2X: 2A = 1 - normalna ženka), 1, 5 - iznad ženke: 2A + 3X (3X: 2A = 1, 5) - bezuspješno). Kod muškaraca je omjer 0, 5 2n = 2A + XY (X: 2A = 0, 5). Sa smanjenjem njegovog pojedinca ostaju mužjaci 3A + XY (X: 3A = 0, 33 - jalovi) - supermeni. Vrijednost koeficijenta između 1 i 0, 5 odgovara fenotipu međuproizvoda pojedinaca prema spolnoj interseksualnosti: 3A + 2X (2X: 3A = 0, 66 - znakovi oba spola su neplodni).
Stoga je bit teorije ravnoteže da ne samo spolni kromosomi, nego i autosomi sudjeluju u određivanju spola. Jedan haploidni skup autosoma govori o pojedinačnim muškim svojstvima. U ovom slučaju, pod određuje se omjerom broja (ravnoteže) autosoma i spolnih kromosoma.
Određivanje spola putem ploidije također se nalazi u pčelama. Ženke su diploidne, a mužjaci su haploidni, jer razviti parthenogenetski od neoplođenih jaja.
Diferencijacija poda u ontogenezi
Počeci gonada u ranim embrijima (do 5. ili 6. tjedna) ne razlikuju se u različitim spolovima i sastoje se od vanjskog sloja - korteksa korteksa i unutarnjeg sloja - medule, ne sadrže stanice zametne linije. Primarne stanice zametne linije nalaze se kod ljudi u trećem tjednu embrionalnog razvoja u ektodermi žumanjčane vrećice. Zatim, pod utjecajem kemotaktičkih signala, migriraju u gonade. Ta je migracija neovisna o spolu. Osnove gonada mogu se razviti u jajnicima ili testisima. Gonadna diferencijacija promatrana je 8. tjedna: na dan 36, testisi počinju lučiti androgene (testosteron), što određuje razvoj muškog spola.
U organizmima koji imaju spolne žlijezde vrši se genetska kontrola stvaranja spolnih karakteristika.
Razlikovanje spola programirano je genetskim skupom spolnih kromosoma koji se formiraju tijekom sjedinjenja ženskih i muških gameta. Genetski spol embrija određen je spolnim kromosomima XX ili XY.
Smjer razvoja spola određen je prisutnošću Y kromosoma. Normalno, X kromosomi sadrže gen za represor (Tfm, gen za feminizaciju testisa) koji sprječava razvoj muškog tipa. Normalni alel gena određuje sintezu proteinskog receptora za androgene, sintetizirane u oba spola. Razvoj muškog fenotipa ovisi o genu Y-kromosoma, HY-antigenu (opisanom u mišu 1955., HA genu). Izlučuju ga primarne muške stanice zametne linije. Y 5NA 0 je odgovoran za proizvodnju testosterona (muški spolni hormoni). Čim te stanice uđu u početke gonada, započinje diferencijacija testisa. HY receptori su prisutni na površini oba tipa gonadnih stanica (odstupanje od razvoja spola kod krava). Smatralo se da je muški fenotip određen cijelim muškim kromosomom. Ali 1990. gena je otkrivena (Sex Region Y), lokalizirana u kariotipu Y kromosoma. U njegovom odsustvu, genotip XY daje ženski fenotip.
RAZVOJ GONADOVA
G ┌┬┬┬┐ │ bakterija Gonad biseksualna
├┼┼┼┤ │ talnaya
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
ako je genotip XX │ │ ako je genotip XY
(7-8 tjedan) ──┴──┐ ┌───┴───┐ (6. tjedan)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
kortikalni ary └┴┘ │ │└┴┴┴┴┴┘│ medularni sloj
sloj ──────┘ └───────┘
(gen definira ženski spol) (gen definira muški spol)
Kod ljudi specifični ženski hormoni koji reguliraju funkcije
ovi znakovi postaju vidljivi, oni su produkt intersticijskog
nas na kraju 7-8 tjedana. Na 20. tkivu jajnika ili sjemene stanice
primarne zametne stanice stanice (Leydigove stanice). Oni tvore
hormonalni seks (testosteron,
gonadama i diferencirani prema estradiolu). 10-12 tjedana za
oogonyev do kraja 2. mjeseca. unutarnji seksualni organizmi
Do kraja trećeg mjeseca u dubini, na 12. tjedan, s izvjesnim
embrioni gonada otkrivaju se na razini androgena u embriju
oociti (profase MI). Počinje diferencijalna maskulinizacija (
ciste jajnika do dobi od 7 mjeseci (srednji uspon) i završava
također će se topiti do 9. mjeseca u jajniku do 20. tjedna.
ima 200-400 tisuća oocita II. U pubertetu, razina est
rogen se povećava i utječe
struktura kostura, kao i
odnosno androgena (prema
ženski i muški tip).
Gonade određuju razvoj primarnih spolnih obilježja i sekundarne. Spolne žlijezde izlučuju hormone (estradiole, androgene), koji zajedno s hormonima endokrinih žlijezda kontroliraju putove diferencijacije spola. Razina hormona zauzvrat kontroliraju geni.
Dakle, proces spolne diferencijacije uključuje:
1) genetska kontrola;
2) regulatorne funkcije hormona.
Postoji teorija djelovanja hormona kao regulatornih čimbenika na gene. Djeluju samo na specifične ciljne stanice. U stanici se proizvodi poseban protein - receptor koji se veže za hormon i istodobno se mijenja, nakon čega dobiva svojstva da inducira rad jednog ili više gena u kromosomima. Kada glukokortikoidi djeluju na stanice maternice, sinteza RNA i proteina se mijenja u njima (vidi dijagram).
Formiranje proteinskih receptora i hormona kontroliraju geni. U slučaju povrede kontrole moguće su anomalije, primjer toga je Morrisov sindrom. Feminizacija testisa (Morrisov sindrom) (Morris, 1953): receptori testosterona su odsutni kod osoba s ovom bolešću. Androgeni se izlučuju u normalnim količinama. U Mauriceovom sindromu, embriogeneza je temelj testisa koji počinju proizvoditi muške spolne hormone. Međutim, takvi embriji ne tvore receptorski protein (recesivna genska mutacija), koji osigurava osjetljivost stanica u razvoju organa na muški spolni hormon. Zbog toga prestaje razvoj muškog tipa i pojavljuje se ženski fenotip. U iznimnim slučajevima moguće je ispraviti takve nedostatke uvođenjem odgovarajućih hormona.
Stoga je izvorna genetska biseksualnost osnova za redefiniranje roda. Muški fetus poprima obilježja ženskog spola. Muški karotip, muški gonad, ženski fenotip. Proporcije tijela su ženske, tu su mliječne žlijezde, skraćena vagina, testisi - u velikoj usnama, u preponskom kanalu, trbušnoj šupljini.
RJEŠENJE KATEGORIJE
Kao posljedica izlaganja hormonima ili kao posljedica patologije receptora ciljne stanice, može doći do redefiniranja spola (Mauriceov sindrom, uklanjanje spolnih žlijezda u pijetlovima).
U prirodi mnogi čimbenici slabe djelovanje gena koji kontroliraju razvoj spola. Primjerice, kod ljudi, u gonadama jednog pojedinca, testisi i jajnici (razvijaju se testisi i jajnici) jednako razvijaju hermafroditizam - fenomen interseksualnosti.
Na temelju kliničkih podataka postoje 3 vrste interseksa:
1) pravi hermafroditizam: prisutnost zametnih stanica oba spola;
2) muški pseudohermafroditizam: postoje samo testisi (testisi, testisi), ženski fenotip;
3) ženski pseudohermafroditizam: postoje samo jajnici, muški fenotip.
Ova se klasifikacija ne podudara s citogenetskim osnovama, jer 46 XY u čovjeku postoje opcije.
Omjer spolova
Primarni omjer spolova (u vrijeme oplodnje trebao bi biti blizu 1: 1, budući da je jednako vjerojatan susret spolnih kromosoma). Tijekom ispitivanja na ljudima, utvrđeno je da se na 100 ženskih zigota formira 140-160 mužjaka. Spermatozoe koje sadrže Y kromosom su lakše, pokretnije i imaju veliki negativni naboj (jaje ima pozitivan naboj). Zbog toga sperma koja sadrži Y opterećuje jaje češće.
Sekundarno - na 100 djevojčica rođeno je 103-105 dječaka. Održivost ženskih gameta, stranih proteina muških embrija. Do 20. godine na 100 djevojaka ima 100 dječaka.
Tercijarno - do 50 godina starosti je 85 muškaraca na 100 žena, a do 85. godine 50 muškaraca na 100 žena. Ženski je organizam više prilagođen, što se, uz ostale razloge, može objasniti i mozaicizmom ženskog organizma na spolnim kromosomima.
M. Loyonova hipoteza o ženskom mozaicizmu na spolne kromosome.
Godine 1949. M. Borr i C. Bertrand otkrili su da je u jezgrama živčanih stanica žena pronađena gomila intenzivno obojenog kromatina. U jezgrama stanica muškarci ga ne otkrivaju. Ta se skupina naziva spolni kromatin (Borrino tijelo) i predstavlja jedan inaktivirani X kromosom.
Na početku razvoja ženskog embrija, oba X-kromosoma funkcioniraju, tj. dvostruko više od muškaraca. To može objasniti veću održivost ženskih zigota.
Godine 1962. M. Lyon postavlja hipotezu o inaktivaciji jednog X kromosoma u ženskom tijelu sisavaca. Kod ženskog embrija, oba kromosoma djeluju do 16. dana embrionalnog razvoja. Na 16. dan, jedan kromosom je inaktiviran formiranjem spolnog kromatina. Ovaj proces je slučajan, tako da oko 1/2 aktivnih stanica održava majčinski X kromosom X 5M 0, a otac je inaktiviran. U drugima, otac je aktivan (X50), a majka je inaktivirana. Ne pojavljuje se ponovna aktivacija. Materinski i paternalni X kromosomi sadrže alelne, ali ne i potpuno identične gene, tj. dominantni alel nalazi se u jednom kromosomu, a drugi je recesivan. Posjedovanje dodatnih gena proširuje adaptivne sposobnosti tijela.
Razine seksualne smjernice seksualne diferencijacije
diferencijacija
┌─┐ ┌┴┐
gamed X └ └─┘ Y
ovum spermatozoon
┌───────────────────┐
genetska hromozomija XX i XY │
└───────────────────┘
┌─────────┴─────────┐
gonadalne ined nedefinirane diferencirane
Gon── gonnye gonade embrija│
│ └───────────────────┘
Ant ant H Y antigen
U razdoblju ┌───────┴───────┐ ┐─────────────
genitalni gen │ │ │1 genitalni
sazrijevanje znakova
│ │ │ │ └────────────┘
─────┴─────┐ ┌─────┴─────┐┐
Week adgondad un adgunad ─────┤ 7-32 tjedna
│ │ jajnici │ testisi ones hormoni andro
│ └───────────┘ └───────────┘┘ │ │ stanični geni
│ │ Leydinga
Y │ pubertet │ │ ┌────────────┐
└───────────────────────────────┘ │ Diferencijacija
hormonalni hormoni hipofize, muški i ženski n živčani živac
Ones hormoni: androgeni (testosteron), estro-putete─ ways
└─ geni (progisteron) │ spol │
┌──────────────┴───────────────┐ └────────────┘
fenotipska │ Sekundarna spolna obilježja priznaki
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
psihološka and spolna i bihevioralna pripadnost
└───────────────────────────────────────────────────┘
Žensko tijelo je otpornije na hladnoću, ionizirajuće zračenje, emocionalna preopterećenja (žene češće plaču, aktivni amini se ističu suzama, zbog čega se krvni tlak smanjuje).
Ako bi hipoteza radila bez ograničenja, ne bi postojale fenotipske razlike između zdravih žena s dva X kromosoma i bolesnika s X 4 0 ili kod muškaraca XY / XXYY. Očito, X kromosom nije potpuno inaktiviran.
Reguliranje omjera spolova.
Ako postavite spermu u konstantno električno polje, doći će do odvajanja X- i Y-kromosoma. Koristi se kod stoke. Moguće je dobiti 80% potomaka željenog spola.
Svijest o pripadnosti muškom ili ženskom spolu sastavni je dio mentalne percepcije spola, mentalnih parametara. Transseksualizam - psihološki hermafroditizam., Fenomen kršenja seksualnog identiteta pojedinca. Slučajevi opsesivne svijesti ljudi koji pripadaju suprotnom spolu poznati su još od antičkih vremena. Dakle, Herodot, u svojoj misteriji "skitske bolesti" opisao je čovjeka koji ne samo da je nosio žensku odjeću, već je usvojio karakterne osobine suprotnog spola. Neki su carevi imali tendenciju prikrivati: na primjer, Caligula. J. D ”Ark.
Razlika između anatomskih i mentalnih parametara je sukob između unutarnjeg okruženja i društvenog. U muškom tijelu živi ženska duša, i obrnuto. Napravite kiruršku korekciju poda. Da biste muškarca pretvorili u ženu, potrebna vam je jedna operacija: vagina se formira iz kože penisa i skrotuma. Od žene da bi muškarac otežao: 3-4 operacije (uklanjanje mliječnih žlijezda, formiranje člana). Ti su ljudi osuđeni na vječne muke: piju hormone, nemaju djece. Institut za ljudsku reprodukciju i planiranje obitelji u Moskvi.
Seksualna diferencijacija uključuje formiranje primarnih (gonadnih) i sekundarnih spolnih karakteristika.
Spol u većini životinja određuje se genetski u vrijeme oplodnje.
X kromosom nosi gen feminizacije testisa (Tfm), normalan
čiji alel određuje sintezu proteinskog receptora za androgene,
koji se sintetiziraju u ženskim i muškim organizmima.
──────┐ - svaki kromosom je nefunkcionalni Tfm - što znači
┌┬┬┐ K┌┬┬┐ varalica razvija korteks.
--────┤ └┴┴┘ ├──────┐┐ XY - sadrži gen I 5HA-gen 0, koji je odgovoran za
Of └──────┘ │ sinteza antigena, koja određuje
│ M = muški = odgovoran za izradu
Ero │ testosteron.
┌──┴──┐ ────┴───┐ Diferencijacija gonada uočena je na 7-10
Of │ │ tjedan života. U 10. tjednu seks može biti op
The │ │ │ za određivanje skupa kromosoma.
└─────┘ └───────┘
Smjer razvoja određen je prisutnošću Y kromosoma. Normalno, XX kromosomi sadrže gen za represor koji sprječava razvoj muškog tipa. Ovaj razvoj ovisi o HY (ON) antigenu, gen I 5HA je odgovoran za proizvodnju testosterona. Nalazi se u nekoliko vrsta (opisano u mišu 1955., Eychwald, Silmser). Izlučuju ga primarne muške stanice zametne linije.
Normalno, kod osoba s kombinacijom spolnih kromosoma, u ženskim genima dominiraju geni koji određuju ženski spol, a prema muškom tipu dominira muški spol.
Genitalije nastaju iz Müller i Wolf kanala, koji potječu iz primarnog bubrega. Kod žena se Müllerovi kanali razvijaju u jajovode i maternicu, a wolfa cijevi atrofiraju. Kod muškaraca, vukovi kanali se razvijaju u sjemene kanale i sjemene mjehuriće. Steroidni hormoni testosteron i dihidrotestosteron sintetizirani su u embrionalnim testisima pod utjecajem hormona majke (horionski gonadotropin). Ovi hormoni djeluju na biopotentne pupoljke.
vanjski i unutarnji genitalni organi: vukove kanale, Mullerovy kanali i urogenitalni sinus. Normalni muški organizam se razvija ako svi ti elementi funkcioniraju. U njihovom nedostatku, oblikuju se ženske spolne karakteristike. Nepotpuni razvoj muškog fenotipa s mužjacima
genotip (muški pseudohermafroditizam).
Suvremena reproduktivna strategija:
- umjetno osjemenjivanje;
- oplodnja in vitro;
- umjetni rast embrija i njegov prijenos u maternicu;
- zamjenska majka.
Većina životinja su dvodomni organizmi. Spol se može promatrati kao skup značajki i struktura koje pružaju način za reprodukciju potomstva i prijenos nasljednih informacija. Spol se najčešće određuje u vrijeme oplodnje, tj. Kariotip zigota igra glavnu ulogu u određivanju spola. Kariotip svakog organizma sadrži kromosome koji su isti u oba spola - autosomi, i kromosomi u kojima se ženski i muški spol međusobno razlikuju - spolni kromosomi. Kod ljudi, "ženski" spolni kromosomi su dva X kromosoma. Tijekom stvaranja gameta, svako jaje dobiva jedan od X kromosoma. Spol u kojem se formiraju gameti istog tipa koji nose X kromosom naziva se homogametic. Kod ljudi je ženski rod homogametičan. "Muški" spolni kromosomi kod ljudi su X kromosom i Y kromosom. Tijekom formiranja gameta, polovica spermatozoida prima X kromosom, druga polovica Y kromosoma. Pod koji ima različite tipove gameta naziva se heterogametni. Kod ljudi je muški spol heterogametan. Ako se formira zigota koja nosi dva X kromosoma, tada se iz nje formira žensko tijelo, ako su X kromosom i Y kromosom muški.
U životinja, sljedeće četiri vrste određivanja kromosomskog spola.
1. Ženski spol je homogametičan (XX), muški spol je heterogametan (XY) (sisavci, osobito čovjek, voćna mušica).
Genetska shema određivanja kromosomskog spola kod ljudi:
Genetska shema određivanja kromosomskog spola u Drosophili:
2. Ženski spol je homogametski (XX), muški spol je heterogametan (X0) (orthoptera).
Genetska shema određivanja kromosomskog spola u pustinjskom skakavcu:
3. Ženski spol je heterogametan (XY), muški spol je homogametski (XX) (ptice, gmazovi).
Genetička shema određivanja kromosomskog spola u goluba:
Yandex.DirectSvi oglasi
4. Ženski rod je heterogametan (X0), muški spol je homogametski (XX) (neke vrste insekata).
Određivanje spola u ljudi se događa mehanizmom XY. Istodobno, heterogametni spol je muški, homogametni - ženski. Određivanje spola je podijeljeno u tri faze: kromosomski, gonadalni i fenotipski.
Dva osnovna pravila za određivanje spola u sisavaca
Klasične embriogenetske studije uspostavile su dva pravila za određivanje spola u sisavaca. Prvi od njih formulirao je šezdesetih godina Alfred Jost na temelju eksperimenata za uklanjanje zametka budućih gonada (gonadalni valjak) u ranim zametcima zeca: uklanjanje valjaka prije nastanka gonada dovelo je do razvoja svih zametaka kao ženki, Predloženo je da se izlučuju gonade muškog efektorskog hormona testosterona, odgovorne za maskulinizaciju fetusa, i predviđa se prisutnost drugog efektora anti-Muller hormona (MIS), koji izravno kontrolira takve anatomske transformacije. Rezultati opažanja formulirani su u pravilu: specijalizacija razvoja gonada u testisima ili jajnicima određuje kasniju spolnu diferencijaciju embrija.
Do 1959. pretpostavljeno je da je broj X kromosoma najvažniji faktor koji kontrolira spol sisavaca. Međutim, otkriće organizama s jednim X kromosomom, koji se razvijaju kao ženke, i pojedinci s jednim Y kromosomom i višestrukim X kromosomima, koji su se razvili kao mužjaci, prisilili su ih da napuste takve pojmove. Formulirano je drugo pravilo za određivanje spola sisavaca: Y-kromosom nosi genetsku informaciju potrebnu za određivanje spola u muškaraca.
Kombinacija gore navedenih pravila ponekad se naziva principom rasta: Kromosomski spol, povezan s prisutnošću ili odsutnošću Y kromosoma, određuje diferencijaciju embrionalnih gonada, što zauzvrat kontrolira fenotipski spol organizma. Takav mehanizam određivanja spola naziva se genetski. GSDi suprotstaviti se takvoj, na temelju kontrolne uloge čimbenika okoliša (eng. ESDili omjer spolnih kromosoma i autosoma (eng. eng.). CSD).
Hormonsko određivanje spola
Rodna definicija može biti predstavljena kao štafeta, koju kromosomski mehanizam prenosi na nediferencirane gonade koje se razvijaju u muške ili ženske spolne organe. Proučavajući ulogu spolnih kromosoma u razvoju gonada, pokazalo se da je prisutnost ili odsutnost Y kromosoma presudna u ljudi. U odsutnosti Y kromosoma javlja se diferencijacija gonada u jajnicima i žena se razvija. U prisutnosti Y kromosoma razvija se muški sustav. Očito, Y kromosom proizvodi tvar koja stimulira diferencijaciju testisa. "Čini se da je osnovni plan prirode bio da se napravi žena, te da dodavanjem Y-kromosoma nastaje čovjek-varijanta." Sljedeća faza releja nastavlja hormone koji određuju proces spolne diferencijacije fetusa i njegov anatomski razvoj. Prilikom rođenja završava se prvi dio programa. Nakon rođenja, palica prelazi na okolišne čimbenike koji završavaju formiranje roda - obično, ali ne uvijek, prema genetskom rodu. Rodno određivanje je složen, višestupanjski proces, koji kod ljudi ovisi, osim biološkog, o psihosocijalnim čimbenicima. To može dovesti do transseksualnog, heteroseksualnog, biseksualnog ili homoseksualnog ponašanja i načina života.
Fiziološke osnove određivanja spola gonada
Fiziološka osnova mehanizma određivanja spola je biseksualnost fetalnih gonada sisavaca. U takvim progonadama, Mullerov kanal i Wolf kanal istovremeno su prisutni - klice genitalnog trakta, odnosno, ženki i mužjaka. Primarno određivanje spola započinje pojavom specijaliziranih staničnih linija u prognozama, Sertoli stanici. U potonjem se sintetizira anti-Müller-ov hormon predviđen Žostom, koji je odgovoran za izravno ili neizravno inhibiranje razvoja Müller-ovog kanala - klica budućih jajovoda i maternice.
Genetski mehanizam za određivanje spola
Humani Y kromosom, koji pokazuje mjesto SRY gena
Godine 1987. David Page i njegovi kolege, ispitujući XX čovjeka koji je naslijedio specifičan fragment Y-kromosoma dužine 280 tisuća nukleotida, i XY ženu s delecijom koja hvata ovu regiju kao rezultat razmjene dijelova između kromosoma. Ovaj fragment prisutan je u Y kromosomu svih stvarnih Eutheria životinja i nalazi se na udaljenosti od 100 tisuća parova baza od granice pseudoautosomalne regije gena ZFY duljine 140 tisuća parova baza.
Homolog ZFY - gen ZFX nalazi se na X kromosomu, a ZFX nije inaktiviran. Oba faktora ZFX i ZFY kodiraju transkripcijske faktore koji sadrže cink prstenaste motive s DNA vezujućom aktivnošću. Daljnja detaljna analiza specifičnih sekvenci Y-kromosoma u osoba s seksualnom inverzijom ograničila je pretraživanje na veličinu od 35 kbp i dovela do otkrića gena koji se smatra istinskim ekvivalentom klasičnog engleskog. Testis odlučujući čimbenik, Taj se gen naziva SRY (eng. Eng. SRY). Određivanje spola Region Y gena).
SRJ koji se nalazi u području određivanja spola i sadrži konzervativnu domenu (HMG-box) koja kodira protein od 80 aminokiselinskih ostataka. Aktivnost SRY gena zabilježena je prije početka perioda diferencijacije trčanja u testis - 10-12 dana mišjeg embrionalnog razvoja i, barem u ovoj fazi, ne ovisi o prisutnosti zametnih stanica. Specifične točkaste mutacije ili delecije u HMG kutiji ovog gena u XY ženama dovode do seksualne inverzije. Prijenos fragmenta DNA od 14 kbp koji sadrži taj gen s bočnim područjima u oplođenu jajnu stanicu homogametičnog pojedinca pomoću mikroinjekcije rezultirao je pojavom mužjaka s kariotipom XX.
SRY funkcije gena
Domena koju kodira HMG kut SRY gena specifično se veže na DNA i uzrokuje njeno savijanje. Savijanje DNA uzrokovano SRY proteinom ili njegovim homolozima koji sadrže HMG domenu mogu se mehanički prenositi na znatnoj udaljenosti i igraju važnu ulogu u regulaciji transkripcije, replikacije i rekombinacije. Područje DNA u kojem je SRY lokalizirano sadrži dva gena koji kodiraju ključne enzime uključene u primarnu gonadnu diferencijaciju muškog tipa: P450 gen aromataze, koji kontrolira konverziju testosterona u estradiol i inhibitor razvoja Miller kanala, koji uzrokuje njihov suprotan razvoj i potiče diferencijaciju testisa ,
Također, SRY genski proizvod sudjeluje u procesima spolne diferencijacije u bliskoj interakciji s drugim genom, nazvanim Z gen, čija je funkcija uobičajeno u inhibiranju specifičnih muških gena. U slučaju normalnog 46XY muškog genotipa, SRY gen kodira protein koji inhibira Z gen, a specifični muški geni su aktivirani. U slučaju normalnog ženskog genotipa 46XX, u kojem je SRY odsutan, Z gen se aktivira i inhibira specifični muški gen, što stvara uvjete za razvoj ženskog tipa.
Kromosomska teorija određivanja spola
Razmotriti određivanje kromosomskog spola. Poznato je da je u dvodomnim organizmima (životinjama i dvodomnim biljkama) omjer spolova obično 1: 1, odnosno, muškarci i žene su jednako uobičajeni. Taj se omjer podudara s razdvajanjem u analizirajućem križu, kada je jedan od križenih oblika heterozigotan (Aa), a drugi je homozigotan za recesivne alele (Aa). U potomstvu u ovom slučaju postoji razdvajanje u odnosu na 1 Aa: 1aa. Ako se rod naslijedi prema istom principu, logično je pretpostaviti da bi jedan spol trebao biti homozigotan, a drugi heterozigotni. Tada bi podjela po spolu trebala biti jednaka 1,1 u svakoj generaciji, što se zapravo i promatra.
Evolucijska teorija roda V. Geodakyan je predložen 1965. Teorija objašnjava s jedinstvenog stajališta mnoge pojave vezane uz spol: spolni dimorfizam u zdravlju i bolesti, omjer spolova, diferencijalna smrtnost i stopa odgovora na spol, uloga spolnih kromosoma i spolni hormoni, asimetrija mozga i ruku, recipročni učinci te psihološke i socijalne razlike među spolovima.
Teorija se temelji na načelu konjugiranih podsustava koji se razvijaju asinkrono. Muškarac je operativan populacijski podsustav, ženski - konzervativan podsustav. Nove informacije iz okoline najprije dosežu muški spol i tek nakon što se mnoge generacije prenesu na ženku, evolucija muškog spola prethodi evoluciji ženke. Ovaj vremenski pomak (dva faza evolucija osobina) stvara dva oblika svojstva (muški i ženski) - spolni dimorfizam u populaciji. Evolucijska "udaljenost" između podsustava potrebna je za pretraživanje i provjeru inovacija.
 Najstrašnija bolest na svijetu
Najstrašnija bolest na svijetu Potpuna i nepotpuna dominacija
Potpuna i nepotpuna dominacija Kako možeš usrećiti svoju djevojku
Kako možeš usrećiti svoju djevojku