Az állatok nemi meghatározásának típusai. Nemek szerinti öröklés
Az ember nemének meghatározása az XY mechanizmus szerint fordul elő. Ebben az esetben a heterogametikus nem férfi, homogametikus nő. A nem meghatározása három szakaszra oszlik: kromoszómális, gonadális és fenotípusos.
Két alapvető szabály az emlősök nemének meghatározására
A klasszikus embriogenetikai vizsgálatok két szabályt hoztak létre az emlősök nemének meghatározására. Az elsőt az 1960-as években Alfred Jost fogalmazta meg a jövőbeni gonidok csírájának eltávolítását (nyálmirigypárna) a nyúl korai embrióiban levő kísérletek alapján: a párnák eltávolítása az ivarmirigy kialakulása előtt az összes embrió nőstény fejlődéséhez vezetett. Azt javasolták, hogy a hímmirigyek szekretálják a magzati masculinizációért felelős effektorhormon tesztoszteront, és megjósolták egy második anti-Muller hormon (MIS) effektor jelenlétét, amely közvetlenül szabályozza az ilyen anatómiai transzformációkat. A megfigyelési eredményeket általában megfogalmazták: a fejléc kialakulásának specializálódása a hereben vagy a petefészekben meghatározza az embrió ezt követő szexuális differenciálódását.
1959-ig az X kromoszómák számát feltételezték a legfontosabb tényezőnek az emlősök nemi kontrolljában. Azonban az egyetlen X kromoszómával rendelkező, nőstényként fejlődő organizmusok felfedezése, valamint hímekként kialakult egy Y kromoszómával és több X kromoszómával rendelkező egyének kénytelenek voltak feladni ezeket az ötleteket. Az emlősök nemének meghatározására szolgáló második szabályt megfogalmazták: Az Y kromoszóma tartalmazza a férfiak nemének meghatározásához szükséges genetikai információkat.
A fenti két szabály kombinációját néha növekedési elvnek hívják: Az Y-kromoszóma jelenlétével vagy hiányával járó kromoszómális nem meghatározza az embrionális gonád differenciálódását, amely viszont a test fenotípusos nemét szabályozza. A nemek meghatározásának hasonló mechanizmusát genetikai jellegűnek hívják. GSD), és ellentétes azzal, amely a környezeti tényezők ellenőrző szerepén alapszik (Eng. ESD) vagy a nemi kromoszómák és az autoszómák aránya CSD).
Hormonális nemi meghatározás
A nemi meghatározást közvetítő verseny formájában reprezentálhatjuk, amelyet a kromoszómamechanizmus megkülönböztethetetlen gonadoknak továbbít, amelyek hím vagy női nemi szervekké alakulnak. A nemi kromoszómáknak a gonidok kialakulásában játszott szerepének vizsgálata során kimutatták, hogy az Y-kromoszóma jelenléte vagy hiánya az embereknél döntő jelentőségű. Az Y kromoszóma hiányában a gonidák differenciálódnak petefészekké és nő alakul ki. Az Y kromoszóma jelenlétében kialakul a férfi rendszer. Nyilvánvaló, hogy az Y kromoszóma anyagot termel, amely serkenti a here differenciálódását. "Úgy tűnik, hogy a természet alapvető célja egy nő készítése volt, és hogy az Y-kromoszóma hozzáadása férfi variációt eredményez." A relé következő szakaszát olyan hormonok folytatják, amelyek meghatározzák a magzat szexuális differenciálódásának folyamatát és annak anatómiai fejlődését. Születéskor a program első része véget ér. Születés után a relé környezeti tényezőkhöz jut, amelyek befejezik a nemek kialakulását - általában, de nem mindig a genetikai nemekkel összhangban. A nemi meghatározás egy összetett többlépcsős folyamat, amely a biológiai mellett a személy pszichoszociális tényezőitől is függ. Ez transzszexualizmushoz, heteroszexuális, biszexuális vagy homoszexuális viselkedéshez és életmódhoz vezethet.
A nemi szájmirigy szintjének fiziológiai alapjai
A nemi meghatározás mechanizmusának élettani alapja az emlős embrionális gonidok biszexualitása. Ilyen futások során a Mueller-csatorna és a Wolf-csatorna egyszerre van jelen - a nemi és a nőstények, illetve a férfiak nemi szakaszai. Az elsődleges nem meghatározása a speciális sejtvonalak megjelenésével kezdődik - a Sertoli sejt. Ez utóbbi a Jost által előrejelzett anti-mulleriai hormont szintetizálja, amely a Muller-vezeték - a jövőbeni petevezeték csíra és a méh - fejlődésének közvetlen vagy közvetett gátlásáért felelős.
A nemi genetikai meghatározás mechanizmusa
Humán Y kromoszóma az SRY gén lokalizációjával
1987-ben David Page és munkatársai feltárják az Y kromoszóma egy adott fragmentumának öröklését, amely 280 ezer nukleotidpár hosszúságú, és egy XY nőstény egy olyan delécióval rendelkezik, amely ezt a régiót megragadja a kromoszómák közötti helycsere eredményeként. Ez a fragmentum jelen van az összes Eutheria állat Y-kromoszómájában, és az ál-autoszomális régió határától 100 ezer nukleotidpár távolságra helyezkedik el. A ZFY gén 140 ezer pár nukleotid hosszúságú.
ZFY homológ - a ZFX gén megtalálható az X kromoszómán, és a ZFX nem kerül inaktiválásra. Mindkét ZFX és ZFY faktor transzkripciós faktorokat kódol, amelyek DNS-kötő aktivitással rendelkező cinkujj-motívumokat tartalmaznak. Az Y-kromoszómák specifikus szekvenciáinak további részletes elemzése a szexuális inverzióval rendelkező egyénekben a kutatást 35 kbp területre korlátozta, és egy olyan gén felfedezéséhez vezetett, amelyet a klasszikus angol valóságnak megfelelőnek tekintnek. A herét meghatározó tényező. Egy ilyen gént SRY-nek hívnak. Nemet meghatározó Y régió gén).
SRY a nemek meghatározásának területén található, és tartalmaz egy konzervatív domént (HMG box), amely 80 aminosavmaradékot tartalmazó fehérjét kódol. Az SRY gén aktivitását megfigyeltük a proguszió herékre történő differenciálódásának kezdete előtt - az egérben az embrionális fejlődés 10-12. Napja, és legalább ebben a szakaszban nem függ a csírasejtek jelenlététől. XY nőkben a gén HMG dobozában lévő specifikus pontmutációk vagy deléciók nemek közötti inverziót eredményeznek. Ezt a gént tartalmazó, körülbelül 14 kbp méretű DNS-fragmentumnak a szomszédos régiókkal történő átvitele egy homogametikus egyed megtermékenyített petesejtjébe mikroinjektálás útján a XX kariotípusú hím megjelenését eredményezte.
SRY gén funkciók
Az SRY gén HMG-box által kódolt domén specifikusan kötődik a DNS-hez és meghajlik. Az SRY protein vagy annak HMG domént tartalmazó homológjai által kiváltott DNS hajlítás jelentős távolságon keresztül mechanikusan átvihető, és fontos szerepet játszhat a transzkripció, replikáció és rekombináció szabályozásában. Az a DNS-régió, amelyben az SRY lokalizálódik, két olyan gént kódol, amelyek kulcsfontosságú enzimeket kódolnak, amelyek részt vesznek az elsődleges gonád differenciálódásában a férfi típus szerint: a P450 aromatáz gén, amely szabályozza a tesztoszteron ösztradiollá történő átalakulását, és az a tényező, amely gátolja a Miller vezetékek fejlődését, ami a fordított fejlődést és elősegíti a herék differenciálódását .
Az SRY gén terméke szintén részt vesz a szexuális differenciálódás folyamatában, szoros kölcsönhatásban egy másik, a Z génnek nevezett génnel, amelynek funkciója általában a specifikus férfi gének elnyomása. A normál 46XY férfi genotípus esetén az SRY gén olyan fehérjét kódol, amely gátolja a Z gént, és specifikus hím gének aktiválódnak. A 46XX normál női genotípus esetén, amelyben nincs SRY, a Z gén aktiválódik és gátolja a specifikus hím gént, ami megteremti a fejlődés feltételeit a nőstípus szerint.
A nemek meghatározásának kromoszómális elmélete
Fontolja meg a kromoszóma neme meghatározását. Ismeretes, hogy a kétéltű szervezetekben (állatok és kétéltű növények) a nemi arány általában 1: 1, azaz a férfi és a nő egyedeket azonos mértékben találják meg. Ez az arány egybeesik az elemzés keresztezésének hasadásával, amikor az egyik keresztezett forma heterozigóta (Aa), és a másik homozigóta a recesszív alléleknél (Aa). Az utódok esetében ebben az esetben megoszlik az 1. viszony Aa: 1aa. Ha a szexet ugyanazon az elv alapján öröklik, akkor logikus lenne azt feltételezni, hogy az egyik nem homozigóta, a másik pedig heterozigóta. Ezután a nemek közötti megoszlásnak minden nemzedékben 1,1-nek kell lennie, amit valójában megfigyelnek.
Evolúciós nemi elmélet az elmélet egyetlen szempontból magyarázza a nemekkel kapcsolatos számos jelenséget: szexuális dimorfizmus a normában és a patológiában, a nemek aránya, a különféle mortalitás és a nemek reakciójának normája, a nemi kromoszómák és nemi hormonok szerepe, az agy és a kéz aszimmetriája, kölcsönös hatások, valamint a nemek közötti pszichológiai és társadalmi különbségek.
Az elmélet az aszinkron módon fejlődő konjugált alrendszerek elvén alapszik. A férfi nem operatív a populáció alrendszere, nő konzervatív alrendszer. A környezetből származó új információk először a férfi nemhez jutnak, és csak sok generáció után kerülnek át a nőkre, ezért a férfi evolúció megelőzi a nő evolúcióját. Ez az időeltolás (kettő fázis a tulajdonság evolúciója) a tulajdonság két formáját (férfi és női) hozza létre - a szexuális dimorfizmus a populációban. Az alrendszerek közötti evolúciós „távolság” szükséges az innovációk kereséséhez és ellenőrzéséhez.
Nem - ez egy szervezet morfológiai, élettani, biokémiai, viselkedési és egyéb jeleinek kombinációja, amelyek meghatározzák a szaporodást.
Azokat a jeleket, amelyek alapján a különféle nemű egyének különböznek, elsődleges és másodlagos jelölésekre osztják. Az elsődleges anyagokat olyan szervezetek képviselik, amelyek biztosítják a ivarsejtek képződését és megtermékenyülését (ivarcsontok, nemi szervek, szervek). Ezek a külső és belső nemi szervek, amelyek az embriogenezisben le vannak fektetve. Másodlagos - nem vesz részt a szexuális szaporodásban. Fejlődik nemi hormonok hatására és pubertáskor jelentkeznek (embereknél 12-15 éves korban). Ezek az izom-csontrendszer fejlődésének jellemzői, a bőr alatti zsír, a haj, a hanghang, a viselkedés; madarakban - ének, tollazat stb.
Az egyén nemekkel kapcsolatos jellemzői három kategóriába sorolhatók:
1) a padlóra korlátozva,
2) nem ellenőrzött
3) a padlóhoz csatolva.
Az előbbi kialakulása mindkét nem autoszómájában található géneknek köszönhető, de csak egy nemben fordul elő. Tehát a tojásrakási gének jelen vannak a csirkékben és a kakasokban, de csak a csirkékben fordulnak elő. A tej és a tejgének a szarvasmarhákon hasonlóan viselkednek. Ezt a jelenséget a megfelelő nemi hormonoknak való kitettség kapcsán lehet megfigyelni.
A második példa a férfiak teheneiben a kürtesség és a nőstényeknél a holtosság megnyilvánulása. Embernél: kopaszság férfiakban, köszvény - férfiakban 80%, nőknél 12%.
Azokat a tulajdonságokat, amelyek fejlődését a nemi kromoszómák génei szabályozzák, nemi kötésnek nevezik. Ilyen jelek kb. 200. Színvak, hemofília kapcsolódik az X kromoszómához; Y-kromoszómával - hipertricózis, ichtiózis.
A padló meghatározásának típusai.
1. Óriási - a megtermékenyítésig. A nemi kromoszómák aránya ebben nem játszik szerepet, mert az oociták diploidok. (Egyes férgek, rotifériák - nőstények fejlődnek a nagy petesejtekből, a férfiak kis petesejtekből).
2. Singamny - a nemek genetikai meghatározása a megtermékenyítés során, amely a nemi kromoszómák kombinációjának jellegétől vagy a nemi kromoszómák és autoszómák arányától függ.
3. Epigám - a külső környezet (bonelia féreg) hatására.
A padló kromoszómális meghatározása
┌─────────────────────────┬───────────────────┬──────────────────┐
│ Kromoszóma típusok │ genotípusok │ ivarsejtek típusai │
│ nemek meghatározása ├─────────┬─────────────────────────────
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
│ férfi heterogenitás │ │
│ Orthoptera rovarok │ │ │ │ │
Prot (Protenor hibák, hibák, │ │ │ │
│ pókok, szöcskék) │ ХО │ ХХ │ Х, О │ Х │
RosDrosophila │ XY │ XX │ X, Y │ X │
│Görénytelen állatok │ │ │ │
(Emlősök, emberek) │ XY │ XX │ X, Y │ X │
│ │ │ │ │ │
│ női heterogenitás │ │
│Ptitsy. hal, pillangók, │ │ │ │ │
Selyemhernyó, hüllő, zem-│ │ │ │ │
│novodnye. │ XX │ XY │ X │ X, Y │
Lepkék és egyéb fordított │ │ │ │ │
Éjszaka │ XX │ XO │ X │ X, O │
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
A kromoszómális nemek genetikai ellenőrzéssel történő meghatározása a szintagmatikus meghatározás típusához tartozik. A szexért felelős személyeket nemi kromoszómáknak nevezzük. Egy normál hím gameta hordozza az X vagy Y kromoszómát, és az összes pete hordozza az X kromoszómát. A meiozis során fellépő normál kromoszóma-divergencia esetén a szokásos X és Y kromoszómakészlettel normál petesejtek és spermiumok alakulnak ki. A zigóta nemét a XX és XY ivarsejtek aránya határozza meg (homogametikus és heterogametikus).
Correns nemi kromoszómális elmélete (1907) szerint a nemet a nemi kromoszómák kombinációja határozza meg a megtermékenyítés során. A kromoszóma nemek meghatározásának következő típusait különböztetjük meg: XY, XO, ZW, ZO.
Ha a mitózis vagy a meiosis folyamata megzavart, gynandomorph egyének alakulhatnak ki. Az ilyen egyének különböző sejtjeiben a nemi kromoszómák tartalma eltérő lehet (mozaik). Drosophila légyben: XX és XO, emberekben XX és XY, amelyekkel kapcsolatban a testrészek különböző nemi jellemzői lehetnek. Lehetnek más mozaicizmus esetei: XX / XXX, XY / XXX; XO / XXY et al.
Ha a nemi kromoszómák nem térnek el egymástól, akkor a nemi kromoszómák 12 lehetséges kombinációja lehet az emberi zigótákban, ez az oka az emberi kromoszóma rendellenességeknek.
│ X │ XX │ Ó
─────┼────────┼─────────┼────────
X., XX., XXX., XO
Y │ XY │ XXY │ YO
XY │ XXY │ XXXY │ XYO
O │ XO │ │
Amennyiben a nemi kromoszómák nem különböznek egymástól a meiosis során, nőstényekben XX és O ivarsejtek alakulnak ki. és XY és O férfiaknál. A megtermékenyítésben való részvételkor a zigóták a nemi kromoszómák szokatlan kombinációjával alakulnak ki. Emberben ilyen rendellenességek előfordulnak 1 600-700 újszülöttnél. A Zygote YO korai stádiumban meghal; Az ХХХ, ХХY, ХО egyének életképesek és nemük az „Y” kromoszóma jelenlététől vagy hiányától függ, amely bármilyen számú X kromoszómával szabályozza a férfi nemi vonások kialakulását, fejlődését és serkenti a herék kialakulását. A túl sok X kromoszóma alkotmányos rendellenességeket és értelmi hibákat okoz. De a természetben vannak olyan egyének, amelyekben az „Y” kromoszóma genetikailag semleges, és nincs különösebb hatással a nem meghatározására.
Drosophila-ban XO típusú egyedeket találtak, amelyek hímek, de termékenyek voltak (1916, K. Bridges), és a XXY. Egyedek normál szaporodású nőstények.
A szex kiegyensúlyozott elmélete (Bridges, 1922). Megvizsgálták a nemi szervek és az autoszómák arányát.
2n kromoszómakészlettel rendelkező normál nőstényeknél az autoszómák és az X-kromoszómák aránya 1: 2n \u003d 2A + 2X (2X: 2A \u003d 1 egy normál nő), 1,5, a nőstény felett: 2A + 3X (3X: 2A \u003d 1,5) -besplodna). Férfiakban ez az arány 0,5 2n \u003d 2A + XY (X: 2A \u003d 0, 5). Az egyének csökkenésével a 3A + XY (X: 3A \u003d 0, 33 - kopár) hímek maradnak. Az 1 és 0, 5 közötti együttható a nemben középszemélyek fenotípusának felel meg: 3A + 2X (2X: 3A \u003d 0, 66 - mindkét nem nevének jelei, sterilek).
Így az egyensúlyelmélet lényege, hogy nemcsak a nemi kromoszómák, hanem az autoszomák is részt vesznek a nemek meghatározásában. Az autoszómák egy haploid halmaza hímeket jelent. Ebben az esetben a nemet az autoszómák és a nemi kromoszómák számának (egyensúlyának) aránya határozza meg.
A nemek ploid általi meghatározása a méhekben is megtalálható. A nőstény méhek diploidok, a hímek pedig haploidok, mert parthenogenetikusan fejlődik ki a megtermékenyítetlen tojásokból.
A padló differenciálódása az ontogenézis során
A korai embriók (az 5. vagy a 6. hétig) csontritkulása nem különbözik a különféle neműek között és a külső rétegből - agykéreg kéregéből és a belső rétegből - a medullaból áll, nem tartalmaz csírasejteket. Az embrionális út elsődleges sejtjeit az embrionális fejlődés harmadik hetében a tojássárgás ektodermájában találják meg az emberekben. Ezután a kemotaktikus szignálok hatására migrálnak az ivarmirigyekbe. Ez a migráció nemtől független. A petefészekben vagy a herékben kialakulhatnak a gonád alulmenetek. A nyálmirigyek differenciálódását a 8. héten figyeljük meg: a 36. napon a herék androgének (tesztoszteron) szekrécióját kezdi, amely meghatározza a férfi nem fejlődését.
A gonidális szervezetekben a nemi jellemzők kialakulásának genetikai ellenőrzését végzik.
A nemi differenciálódást a nemi kromoszómák genetikai halmaza programozza, amely a női és férfi ivarsejtek fúziója során alakul ki. Az embrió genetikai nemét a XX vagy XY nemi kromoszóma határozza meg.
A nemi fejlődés irányát az Y kromoszóma jelenléte határozza meg. Általában az X kromoszómák tartalmaznak egy represszor gént (Tfm here-feminizációs gén), amely megakadályozza a férfi típus kialakulását. A normál gén allél meghatározza mindkét nemben szintetizált androgének proteinreceptorjának szintézisét. A hím fenotípus szerinti fejlődés az Y-kromoszóma géntől, a H-y antigéntől függ (egerekben 1955-ben leírták; HA gén). Az elsődleges hím csírasejtsejtek szekretálják. Az Y 5NA 0 felelős a tesztoszteron (férfi nemi hormon) termeléséért. Amint ezek a sejtek belépnek a gonidok alsó részébe, megkezdődik a herék differenciálódása. A H-Y receptorok mindkét típusú gonádsejtek felületén vannak (eltérés a nemek fejlődésétől a teheneknél). Úgy véljük, hogy a hím fenotípusát a teljes férfi kromoszóma határozza meg. De 1990-ben felfedezték egy gént (Y szex régió), amely az Y kromoszóma kariotípusában található. Ennek hiányában az XY genotípus női fenotípust ad.
A GONAD FEJLESZTÉSE
│ ┌┬┬┬┐ b Az embrió biszexuális gonádja
│ ├┼┼┼┤ │ al
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
ha a XX genotípus │ │, ha a XX genotípus
(7-8 hét) ┌──┴──┐ ┌───┴───┐ (6. hét)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
kortikális │ └┴┘ │ │└┴┴┴┴┴┘│ meduláris réteg
réteg └─────┘ └───────┘
(a gén határozza meg a női nemet) (a gén határozza meg a férfi nemet)
Az embernek vannak specifikus női hormonjai, amelyek szabályozzák a
a jelek észrevehetők, az intersticiális termékek termékei
minket a 7-8 hét végén. A petefészek vagy petesejtek 20. szövetén
elsődleges csírasejtek, beleértve (Leydig-sejteket). Ezek alkotnak
hormonális szex (tesztoszteron,
gonadok és differenciálódnak ösztradiolra). 10–12 hétnél-
ovogony a 2. hónap végére. belső nemi szervek
A 3. hónap végére bennünk a mélységben, a 12. héten egy bizonyos időben
az embriómirigyeket az embrió androgének szintjén detektálják
petesejtek (MI fázis). Differ - kezdődik a masculinization
petefészek keringés a középső réteg 7. hónapjáig) és befejezi-
a petefészek 9. hónapjáig a 20. hétig megolvad.
200-400 ezer oocita van II. A pubertás idején az est
a rogen növekszik és befolyásolja
a csontváz szerkezete, valamint a
androgének, ill
női és férfi típus).
A gonondok meghatározzák az elsődleges szexuális és másodlagos jellemzőket. A nemi mirigyek hormonokat választanak ki (ösztradiolok, androgének), amelyek az endokrin mirigyek hormonjával együtt szabályozzák a nemek differenciálódását. A hormonszintet viszont a gének szabályozzák.
Így a szexuális megkülönböztetés folyamata magában foglalja:
1) genetikai védekezés;
2) a hormonok szabályozó funkciói.
Van egy elmélet a hormonok mint gének szabályozó tényezőinek hatásáról. Csak meghatározott célsejtekre hatnak. A sejtben egy speciális fehérje termelődik - egy olyan receptor, amely kötődik a hormonhoz, és ezzel egyidejűleg megváltozik, ezután megszerezheti a tulajdonságokat, hogy egy vagy több gén működését indukálja a kromoszómákban. A méhsejtekre adott glükokortikoidok hatására megváltozik az RNS és fehérjék szintézise (lásd. Séma).
A receptor fehérjék és hormonok képződését a gének szabályozzák. Kontroll megsértése esetén rendellenességek léphetnek fel, amelyek példája a Morris-szindróma. Tetuláris feminizáció (Morris-szindróma) (Morris, 1953): E betegségben szenvedő egyének hiányzik a tesztoszteron receptorokból. Az androgének normál mennyiségben választódnak ki. A Moris-szindróma során az embriógenezist olyan herék fektetése kíséri, amelyek megkezdi a férfi nemi hormonok termelését. Ilyen embriókban azonban nem alakul ki receptor protein (recesszív génmutáció), amely biztosítja a fejlődő szervek sejtjeinek érzékenységét a férfi nemi hormonra. Ennek eredményeként a férfi típus fejlődése megáll és a női fenotípus megnyilvánul. Kivételes esetekben ezeket a hibákat megfelelő hormonok bevitelével lehet kijavítani.
Így az ős genetikai biszexualitás a nemek újradefiniálásának alapja. A férfi magzat megszerezte a női nemre jellemző tulajdonságokat. Férfi kariotípus, hímmirigyek, női fenotípus. A testarány nőstény, vannak emlőmirigyek, rövidített hüvely, a herék a labia majora-ban, az inguinalis csatorna és a hasi üreg.
A padló újrafogalmazása
Hormonoknak való kitettség vagy a célsejt receptorának patológiája eredményeként megváltozhat a nemek újradefiniálása (Moris-szindróma, a nemi mirigyek eltávolítása a kakasokban).
A természetben számos tényező gyengíti a nemek fejlődését szabályozó gének hatását. Például, ha az egyén egyénének gondekában egyaránt kifejlődik a here és a petefészke (a herék és a petefészek kialakul), a hermaphroditizmus az interszexualitás jelensége.
A klinikai adatok alapján az intersex 3 típusát különböztetik meg:
1) valódi hermaphroditizmus: mindkét nemben csírasejtek jelenléte;
2) férfi pszeudohermaphroditizmus: csak herék vannak (herék, herék), női fenotípus;
3) női pszeudohermaphroditizmus: csak petefészek található, a férfi fenotípus.
Ez a besorolás nem esik egybe a citogenetikai alapokkal, tehát 46 XY egy embernél van lehetőségek.
Nemek aránya.
Az elsődleges nemi arány (a megtermékenyítés idején 1: 1-nek kell lennie, mivel a nemi kromoszómák találkozása szintén valószínű). Egy emberben végzett vizsgálat feltárta, hogy 100 nőstény zigótán 140-160 hím cigota képződik. Az Y kromoszómát tartalmazó sperma könnyebb, mozgathatóbb és nagy negatív töltésű (a tojás pozitív töltéssel rendelkezik). Ezért az Y-tartalmú sperma gyakran megtermékenyíti a petesejét.
Másodlagos - a születés időpontjáig 100 lányonként 103-105 fiú született. A női ivarsejtek életképessége, a férfi embriók proteinjeinek idegessége. 20 éves korig 100 lányonként 100 fiú van.
Felsőfokú - 50 éves korig 100 nőnként 85 férfi van, 85 éves korukra pedig 100 férfi 50 férfi. A női test jobban alkalmazkodik, ami más okokkal magyarázható a női test nemi kromoszómák általi mozaikszerűségével.
Hipotézis M. Loyon a női mozaicizmusról a nemi kromoszómákban.
1949-ben M. Borr és C. Bertrand megállapította, hogy az intenzíven festett kromatin blokkja található a nő idegsejtjeiben. A férfiak sejtjeiben nem észlelhető. Ezt a csomót nemi kromatinnak (Borr test) nevezik, és egy inaktivált X kromoszómát képvisel.
A fejlődés kezdetén mindkét X kromoszóma a női magzatban működik, azaz kettővel több, mint az ember géneinél. Ez megmagyarázza a női zigóták nagy életképességét.
1962-ben M. Lyon hipotézisként felvette egy X kromoszóma inaktiválását a női emlős szervezetben. Egy női magzatban mindkét kromoszóma az embrionális fejlődés 16 napjáig működik. A 16. napon egy kromoszóma inaktiválódik a nemi kromatin képződésével. Ez a folyamat véletlenszerű, tehát körülbelül 1/2 sejtben az anyai X 5M 0 kromoszóma aktív marad, és az apák inaktiválódnak. Másokban az apaság aktív (X 5 O 0), az anyai inaktiválódnak. Újraaktiválás nem történik meg. Az anyai és apai X kromoszómák allél, de nem feltétlenül azonos géneket tartalmaznak, azaz a domináns allél az egyik kromoszómán, a recesszív a másikon található. További gének birtoklása kibővíti a test adaptív képességeit.
Szexuális szintek A szexuális differenciálódás diagnosztikája
különbségtétel
┌─┐ ┌┴┐
gamet X └┬┘ └─┘ Y
ovule─┼─ sperma
┌───────────────────┐
genetikai XX XX és XY kromoszóma │
└───────────────────┘
┌─────────┴─────────┐
ízületi │ megkülönböztethetetlen--
│ et magzati nyálmirigyek │
│ └───────────────────┘
│ │ H-Y antigén
│ ┌───────┴────────┐ ┌────────────┐ időszakban
nemi │ │ │1 nemi │
érés ┌─│ │ │ ┌─│ jelek │
│ │ │ │ └────────────┘
Uter ┌─────┴─────┐ ┌─────────── ┐ │ in utero
│gonad │gonad ├────┤ 7-32 week
│ │ petefészek, │ tes herék és androhormonok
│ │ └───────────┘ └───────────┘ │ sejtgének
│ │ Vezető
│ y pubertás │ │ ┌────────────┐
│ └─────────────────────────────── │ │ │ differenciálás │
hormonális │ hipofízis hormonok, férfi és női │ │ idegek │
Es hormonok: androgének (tesztoszteron), ösztrogén módon │
└─ gének (progeszteron) │ nemek │
┌──────────────┴───────────────┐ └────────────┘
fenotípusos │ Másodlagos szexuális jellemzők │
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
pszichológiai │ nem és magatartás │
└───────────────────────────────────────────────────┘
A női test jobban ellenáll a hidegnek, az ionizáló sugárzásnak, az érzelmi túlterhelésnek (A nők gyakrabban sírnak, aktív aminok szabadulnak fel a könnyekkel, ennek eredményeként csökken a vérnyomás).
Ha a hipotézis korlátozás nélkül működne, akkor nem lenne fenotípusos különbség a két X kromoszómával rendelkező egészséges nők és az X 4 0 betegek, illetve a férfiak XY / XXYY között. Nyilvánvaló, hogy az X kromoszóma nincs teljesen inaktiválva.
A nemek arányának szabályozása.
Ha a spermat állandó elektromos mezőbe helyezzük, az X - és Y kromoszóma elválasztódik. Állattenyésztésben használják. A megfelelő nem leszármazottainak 80% -át megszerezheti.
A férfi vagy női nem ismerete a nemek mentális észlelésének és a mentális paraméterek szerves része. Transzszexualizmus - pszichológiai hermafroditizmus., A személy szexuális identitásának megsértésének jelensége. Az ellenkező nemhez való tartozás kényszeres tudatosságának eseteit az ókortól kezdve ismerték. Tehát Herodotus „Szkíta betegsége” misztériumában egy embert írt le, aki nem csak női ruhát visel, hanem az ellenkező nemű karakterisztikáját is átvette. Néhány császár hajlandó volt öltözködni: például Caligula. J. D ”Arc.
Az anatómiai és mentális paraméterek eltérése a belső környezet és a társadalmi konfliktusok összeférhetetlensége. A női lélek a férfi testben él és fordítva. A műtéti szexuális korrekció megtörtént. Egy nő nővé tételéhez 1 műveletre van szüksége: a pénisz és a herezacskó bőréből hüvely alakul ki. Nehezebb egy férfit nőből előállítani: 3-4 műtét (tejmirigyek eltávolítása, péniszképződés). Ezek az emberek örök gyötrelmekre vannak ítélve: hormonális gyógyszereket fogyasztanak, nincsenek gyermekeik. Az emberi szaporodás és a családtervezés intézete Moszkvában.
A szexuális megkülönböztetés magában foglalja az elsődleges (gonidák) és a szekunder szexuális jellemzők kialakulását.
A legtöbb állat nemét genetikailag a megtermékenyítés időpontjában határozzák meg.
Az X kromoszóma héjas feminizációs gént (Tfm) hordoz, normál
amelynek alléle határozza meg az androgének proteinreceptorjának szintézisét,
amelyek mind a nőstényekben, mind a férfiakban szintetizálódnak.
┌──────┐ - minden kromoszóma nem senegén Tfm -
│ │K│ csalókéreg kéreg kialakulását fejleszti ki.
┌────┤ └┴┴┘ ├──────┐ XY - tartalmazza az I gént, az 5. DNS-t, amely felelős azért
│ └──────┘ │ antigén szintézis meghatározása
│ M │ férfi \u003d a termelésért felelős
Tesztoszteron.
Gon of A gonidok differenciálódását 7-10-kor figyeljük meg
Life life life az élet hete. A 10. héten a padló op
│ │ │ │ Rendezés a kromoszómakészlet alapján.
└─────┘ └───────┘
A fejlődés irányát az Y kromoszóma jelenléte határozza meg. Általában a XX kromoszómák represszor gént tartalmaznak, amely megakadályozza a férfi típus kialakulását. Ez a fejlődés az N-Y (HA) antigéntől függ, az I 5HA gén felelős a tesztoszteron termeléséért. Több fajban található meg (egerekben írták le 1955-ben, Eichwald, Silmser). Az elsődleges hím csírasejtsejtek szekretálják.
Általában a nemi kromoszóma kombinációjával rendelkező egyéneknél a női típusban a női nemet meghatározó gének dominálnak, a férfi - a férfi nemben.
A nemi szervek a mulleri és a Wolf vezetékéből alakulnak ki, amelyek az elsődleges veseből származnak. A nőkben a Müller-vezetékek a petevezetékekbe és a méhbe fejlődnek, míg a Wolf-vezetékek atrófiává alakulnak. A férfiakban a Farkas vezetékek fejlődnek a petevezetékekké és a vezikulákké. Az embrionális herékben a szteroid hormonokat, a tesztoszteront és az 5-dihidrotestoszteront az anyahormon (koriongonadotropin) hatására szintetizálják. Ezek a hormonok a biopotenciális rügyekre hatnak
külső és belső nemi szervek: farkascsatornák, mulleri csatornák és urogenitális sinus. Normális férfi test alakul ki, ha ezen elemek mindegyike működik. Ezek hiányában nők szexuális tulajdonságai alakulnak ki. A hím fenotípusának hiányos kialakulása a hímnél
genotípus (férfi pszeudohermafroditizmus).
Modern reprodukciós stratégia:
- mesterséges megtermékenyítés;
- in vitro megtermékenyítés;
- az embrió mesterséges termesztése és a méhbe történő átültetése;
- helyettes anya.
A legtöbb állat kettős élőlény. A szex olyan tulajdonságok és struktúrák kombinációjának tekinthető, amelyek az utódok reprodukálásának és az örökletes információk továbbításának a módját biztosítják. A nemet leggyakrabban a megtermékenyítéskor határozzák meg, vagyis a zigóta kariotípus fontos szerepet játszik a nem meghatározásában. Az egyes szervezetek kariotípusa mindkét nemben azonos kromoszómákat tartalmaz - autoszómákat, és azokat a kromoszómokat, amelyek alapján a női és a férfi nem különbözik egymástól - a nemi kromoszómákat. Az emberekben a „női” nemi kromoszóma két X kromoszóma. Amikor ivarsejtek képződnek, minden pete megkapja az X kromoszóma egyikét. Egy olyan nem, amelyben azonos típusú ivarsejtek képződnek, és amelyek hordozzák az X kromoszómát, homogametikusnak nevezik. Az emberekben a női nem homogametikus. Az emberben a „férfi” nemi kromoszómák az X és az Y kromoszóma. Amikor ivarsejtek képződnek, a sperma fele megkapja az X kromoszómát, a másik fele az Y kromoszómát. A nemet, amelyben különféle ivarsejtek képződnek, heterogametikusnak nevezik. Az emberekben a férfi nem heterogametikus. Ha olyan zigóta képződik, amely két X-kromoszómát hordoz, akkor nőstény szervezet alakul ki belőle, ha az X-kromoszóma és az Y-kromoszóma hím.
Állatokban a következő négyféle kromoszóma nem meghatározása.
1. A női nem homogén (XX), a férfi heterogametikus (XY) (emlősök, különösen emberek, Drosophila).
A kromoszóma neme meghatározásának genetikai sémája emberben:
A kromoszóma neme meghatározásának genetikai sémája Drosophila-ban:
2. A női nem homogén (XX), a férfi heterogametikus (X0) (orthoptera).
A sivatagi sáskák kromoszóma neme meghatározásának genetikai sémája:
3. A női nem heterogametikus (ХY), a hím homogén (XX) (madarak, hüllők).
A galambban a kromoszóma nemének meghatározása:
Yandex.DirectAll hirdetések
4. A női nem heterogametikus (X0), a férfi egynemű (XX) (egyes rovarok).
Az állatok túlnyomó többségét két nem - egyedülálló hímek és nők - képviselik. Néhány növényfajban két nemű egyedeket is megfigyelnek (kender, máj spherocarpus moha, álmosság stb.). Az egyén nemét meghatározó mechanizmus egyértelmûvé vált az öröklõdés kromoszómális elméletének megalkotása után, bár a nemi öröklõdés törvényeivel kapcsolatban néhány megfigyelés sokkal korábban történt. Régóta ismert, hogy egy faj populációjában átlagosan a férfiak és nők aránya azonos, azaz 1: 1. G. Mendelben ez az arány analógiát váltott ki a monohibrid elemző keresztekkel, amint azt fentebb leírtuk a keresztek elemzésekor, 1: 1 hasítás történik, ha az egyik szülő heterozigóta a vizsgált génnél (Aa),és a másik recesszív homozigóta (Aa).A nemek szerinti megoszlásnál megfigyelt 1: 1 arány lehetővé tette annak feltételezését, hogy az egyik nem heterozigóta, a másik pedig a test nemét meghatározó tényezőhöz homozigóta. Ezt a feltételezést a század elején megerősítették a genetikai kísérletek eredményei, és citológiai megerősítést kaptak a nemi kromoszómák felfedezése után. Az állatok szaporodási sejtjeinek és testsejtjeinek kromoszómáinak szerkezetének tanulmányozása során kiderült, hogy a férfiak és nők eltérőek a kromoszómakészletben. A nemi kromoszómákat W. Wilson és C. Mack Klang vizsgálta részletesen.
Nemi kromoszómák. Kényelmes mérlegelni a Drosophila kromoszómákban a nemek meghatározására szolgáló kromoszóma-mechanizmus jelenlegi gondolatát. Ábrán A 84. ábra egy Drosophila kromoszómakészlet vázlatos ábrázolása, amely mindössze nyolc 4 párt tartalmazó kromoszómából áll. Három párban a kromoszómák homológok egymással és morfológiailag nem különböznek egymástól. A negyedik pár kromoszómái nagyon különbözőek. Az egyik, az úgynevezett X-kromoszóma, rúd alakú, a második (Y-kromoszóma) kampó alakja van, amelyet e kromoszóma egyenlőtlen karjai képeznek.

egy X és egy Y kromoszóma. A kromoszómák egy párját, amelyben a férfiak és a nők különböznek, nemi kromoszómának vagy heterokromoszómának nevezzük, szemben más autoszómáknak nevezett kromoszómákkal (néha A rövidítéssel).
Homo és heterogametikus szex. Drosophila-ban az azonos nemű kromoszómákkal rendelkező nők (XX), a meiosisból származó összes csírasejt azonos nemű kromoszómával rendelkezik (X), amelynek eredményeként az összes ivarsejt azonos. A Drosophila női nemét, amely egyetlen típusú ivarsejtet termelő, homogametikusnak nevezik. A Drosophila hímek viszont kétféle ivarsejt termelnek, amelyek egyikében a nemi kromoszómát az X kromoszóma, a másikban az Y kromoszóma képviseli. A Drosophila-ban a férfi nem heterogametikus.
A nőstényeknek a homo- vagy heterogametikus nemhez való kapcsolódásától függően az összes állatot két csoportra osztják. Az emlősökben, férgekben, rákfélékben, a legtöbb rovarban (beleértve a Drosophila-t), a legtöbb kétéltűben és egyes halakban a férfi nem heterogametikus, a nőstény pedig homogametikus.
Ebbe a csoportba az emberek tartoznak. Az emberi szomatikus sejtek 44 autoszomális kromoszómát tartalmaznak, emellett a nőknek két X kromoszóma van, a férfiaknak pedig egy X kromoszóma és egy Y kromoszóma. A női emberi test kromoszómaképlete 44A + lehet XX, férfi - 44A + XU. Madarakban, hüllőkben, kétéltűekben és halakban egyes rovarok (pillangók és caddis legyek) nőivarúak. Ebben az esetben más szimbólumokat használunk a nemi kromoszómák megjelölésére. Női heterogameticitás esetén a nemi kromoszómái jelzik ZW, és férfi nemi kromoszómák, ZZ. Például azokban a csirkékben, amelyekben a szomatikus sejtekben 78 kromoszóma van, a hím kromoszóma képlete -76A + ZZnő, 76A + ZW.
Egyes fajoknál az evolúciós folyamat során a heterogametikus nem egyik nemi kromoszóma elveszett, amelynek eredményeként kromoszómakészletükben csak egy nemi kromoszóma található. A nemi kromoszóma formulát egy ilyen esetben heterogametikus nemben a következővel jelöljük XOvagyZO.
A padló felosztásának mechanizmusa 1: 1. A nemek meghatározásának kromoszómamechanizmusának ismerete lehetővé teszi a férfi és női egyedek megjelenésének okainak 1: 1 arányú magyarázatát. Mint fentebb megjegyeztük, a homogametikus nem azonos típusú ivarsejteket alkot, amelyek mindegyike azonos nemű kromoszómát hordoz. A heterogametikus nemi ivarsejtekben viszont az egyik vagy a másik nem kromoszóma van jelen. Például Drosophila-ban a nőstények összes csírasejtje egy X kromoszómával rendelkezik, a hímben lévő ivarsejtek felében X kromoszóma, a másik felében Y kromoszóma van.
Ha a tojás (X) spermával megtermékenyítik az X kromoszómával, a két X kromoszóma kombinációja a zigótában nőstény képződést eredményez (XX). Ugyanazon petesejt spermával történő megtermékenyítése az U kromoszómával hím (XU) megjelenését okozza. Mivel az egyik vagy másik sperma megtermékenyítésében való részvétel ugyanolyan valószínű, a férfiak és nők megjelenése azonos eséllyel jár. A különféle fajú állatokban nagyszámú újszülött egyén esetében a nemek aránya 1: 1 vagy nagyon közel áll. Tehát a 100 újszülött közül hímekben 51 ember van, 51 kutyában, 49 csirkében, 50 egérben, 49 juhban, 51 szarvasmarhaban és 52. lovakban. Ezek a számok jellemzik az egyének arányát különböző neműek születésekor. A jövőben ezek az arányok nagymértékben változhatnak, különféle nemű egyének egyenlőtlen túlélése miatt. Tehát az emberekben 50 éves korig a férfiak és a nők aránya 85: 100, 85 éves korukra pedig 50: 100.
Az egyik vagy másik nem nem a kromoszómamechanizmus által okozott megnyilvánulása bizonyos esetekben kísérletileg megváltoztatható, amint azt csirkék, selyemhernyók stb. Mutatják. A két- és többnemzetiségű, hím és nőstény növényekben a nemi kromoszómákat is megvizsgálták; e növények többségében heterogametikus a férfi nem.
A nemek genetikája
A nemre karakterkomplex tartozik, amelyet a kromoszómákon elhelyezkedő gének határoznak meg. Az emberi test sejtjeiben a kromoszómák párosított diploid halmazok. A kétéltű egyéneknél a férfiak és nők kromoszóma-komplexe nem azonos, és kromoszómák (nemi kromoszómák) szerint különbözik egymástól. A pár ugyanazon kromoszómáit X (X) kromoszómának hívták, pár nélkül, a másik nemben hiányzik - az U (játék) kromoszómának; a többi, amelyben nincsenek különbségek, autoszómák (A).
Egy nő sejtjei két azonos nemű kromoszómát tartalmaznak, amelyeket XX jelöléssel látnak el, a férfiakban két páratlan X és Y kromoszóma képviseli őket. Így a férfi és a nő kromoszómakészlete csak egy kromoszómában különbözik egymástól: egy nő kromoszómakészlete 44 autoszómát tartalmaz + XX, férfi - 44 autoszómát + XY.
Az emberi csírasejtek megosztása és érése során haploid számú kromoszómával rendelkező ivarsejtek alakulnak ki: a tojások általában 22 + X kromoszómát tartalmaznak. Így nőkben csak egyfajta ivarsejt képződik (ivarsejtek az X kromoszómával). A férfiakban a ivarsejtek 22 + X vagy 22 + Y kromoszómát tartalmaznak, és kétféle ivarsejt képződik (ivarta X-kromoszómával és ivarta Y-kromoszómával). Ha az X kromoszómával rendelkező sperma bejut a petesejtbe a megtermékenyítés során, nőstény magzat képződik, hím pedig az Y kromoszómával.
Ezért az ember nemének meghatározása függ a férj csírasejtekben - spermákban, amelyek megtermékenyítik a petesejtet, X- vagy Y-kromoszómákban - való jelenléttől.
A kromoszóma neme meghatározásának négy fő típusa van:
1. A férfi nem heterogén; A ivarsejtek 50% -a hordozza az X-, 50% -U - kromoszómát, például emberek, emlősök, dipteranok, bogarak, bogarak (4. dia).
2. A férfi nem heterogén; A ivarsejtek 50% -a hordoz X-, 50% - nem rendelkezik nemi kromoszómával, például szöcske, kenguru (7. dia).
3. A női nem heterogén; A ivarsejtek 50% -a hordozza az X-, 50% -a ivarsejteket - Y– kromoszómát, például madarak, hüllők, farkú kétéltűek, selyemhernyó (7. dia).
4. A női nem heterogén; A ivarsejtek 50% -a hordoz X-, 50% -uk nem rendelkezik nemi kromoszómával, például egy anyajegy.
Azon tulajdonságok öröklődését, amelyek génei a nemi kromoszómán helyezkednek el, öröklésnek nevezzük, össze van kötve a padlóval.
26. A genotípus mint egy integrált rendszer. A gének kölcsönhatása, a gének sokféle hatása.
A genotípus mint holisztikus rendszer
A gének tulajdonságai. Alapján keresek Példákkal a karakterek öröklődéséről a mono- és dihibrid keresztekben azt a benyomást keltheti, hogy a szervezet genotípusa különálló, egymástól függetlenül működő gének összegéből áll, amelyek mindegyike csak saját vonása vagy tulajdonsága fejlődését határozza meg. Ez a gondolat, hogy egy gén egy vonással közvetlenül és egyértelműen kapcsolódik, általában nem igaz. Valójában óriási számú élő organizmus tulajdonsága és tulajdonsága van, amelyeket két vagy több génpár határoz meg, és fordítva: egy gén gyakran sok tulajdonságot irányít. Ezenkívül egy gén hatását megváltoztathatja más gének közelsége és a környezeti feltételek. Tehát a ontogenezis során nem az egyes gének működnek, hanem a teljes genotípus integrált rendszerként, komplex kötésekkel és kölcsönhatásokkal az alkotóelemek között. Ez a rendszer dinamikus: új allélek vagy gének megjelenése mutációk eredményeként, új kromoszómák és még új genomok kialakulása a genotípus észrevehető változásához vezet az idő múlásával.
A gén hatásának a genotípus, mint rendszer összetételében történő megnyilvánulásának jellege különböző helyzetekben és körülmények között változhat befolyása különböző tényezők. Ez könnyen látható, ha figyelembe vesszük a gének tulajdonságait és a jelekben történő megnyilvánulásuk jellemzőit:
Egy gén működése diszkrét, azaz aktivitása során elkülönül más génektől.
A gén specifikus a megnyilvánulásában, azaz felelős a szervezet szigorúan meghatározott tulajdonságáért vagy tulajdonságaiért.
Egy gén fokozatosan hathat, vagyis fokozhatja a tulajdonság megnyilvánulásának mértékét a domináns allélek számának (géndózis) növekedésével.
Egyetlen gén befolyásolhatja a különféle tulajdonságok kialakulását - ez a gén többszörös vagy pleiotróp hatása.
A különböző gének ugyanolyan hatással lehetnek ugyanazon tulajdonság kialakulására (gyakran kvantitatív tulajdonságok) - ezek több gén vagy poligén.
Egy gén kölcsönhatásba léphet más génekkel, ami új tulajdonságok megjelenéséhez vezet. Az ilyen interakciót közvetett módon hajtják végre - reakcióik termékein keresztül, amelyeket ellenőrzésük alatt szintetizálnak.
A gén működését módosíthatjuk annak helyét a kromoszómán (pozícióhatás) vagy különféle környezeti tényezők hatására.
Allerikus gén kölcsönhatások. Ezt a jelenséget, amikor több gén (allél) felelős egy tulajdonságért, gének interakciójának nevezzük. Ha ezek ugyanazon gén allélei, akkor az ilyen interakciókat nevezzük allél és különböző gének alléljai esetén - nemallelikus.
Az allél kölcsönhatások következő fő típusait különböztetjük meg: dominancia, hiányos dominancia, túlsúly és kódolás.
uralom - az egyik gén két alléljának kölcsönhatásának típusa, amikor egyikük teljesen kizárja a másik működésének megnyilvánulását. Ez a jelenség a következő körülmények között lehetséges: 1) egy heterozigóta állapotban lévő domináns allél olyan termékek szintézisét biztosítja, amely elegendő olyan tulajdonság megjelenéséhez, amely azonos minőségű, mint a domináns homozigóta szülő forma állapotában; 2) a recesszív allél teljesen inaktív, vagy aktivitásának termékei nem lépnek kölcsönhatásba a domináns allél aktivitásának termékeivel.
Az allél gének ilyen interakcióira példa a lila dominancia. szín borsóvirág fehér, sima alakú mag felett, ráncos, sötét haj világosság felett, barna szem kék felett embernél stb.
Hiányos dominancia vagy az öröklés közbenső jellege, megfigyelhető abban az esetben, ha a hibrid (heterozigóták) fenotípusa különbözik mindkét szülő homozigóta fenotípusától, azaz a tulajdonság kifejeződése közbülső, nagyobb vagy kisebb eltérést mutat az egyik vagy a másik szülő felé. Ennek a jelenségnek az a mechanizmusa, hogy a recesszív allél inaktív, és a domináns allél aktivitásának mértéke nem elegendő a domináns tulajdonság kívánt megnyilvánulási szintjének biztosításához.
A hiányos dominancia például az öröklés. szín virágok éjszakai szépségű növényekben (3.5. ábra). Amint az a diagramból látható, a homozigóta növények vagy vörös színűek (AA) akár fehér (Aa) virágok és heterozigóta (Aa) - rózsaszín. Ha a növényeket piros virágokkal és fehér virágokkal keresztezik az F1-ben, akkor minden növénynek rózsaszínű virágai vannak, azaz megfigyelték az öröklés közbenső jellege. Hibridek keresztezésekor a virágok rózsaszín színezése F 2 véletlen egybeesés van a hasítás fenotípus és genotípus alapján, mivel a domináns homozigóta (AA) eltér a heterozigótától (Aa). Tehát a vizsgált példában az éjszakai szépség növényeivel szétválasztva F 2 a virágok színe általában a következő - 1 piros (AA): 2 rózsaszín (Aa): 1 fehér (Aa).
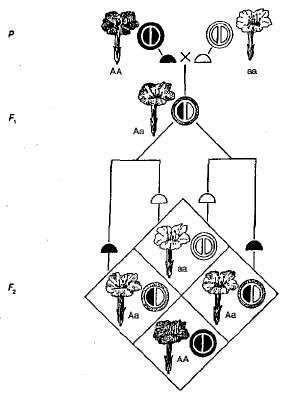
Ábra. 3.5. Az éjszakai szépségben hiányos uralkodó virágok színének öröklődése.
A hiányos dominancia széles körben elterjedtnek bizonyult. Megfigyelhető az ember göndör szőrének öröklésében, a szarvasmarhák színében, a tollazat színezésében a csirkékben, valamint sok más morfológiai és élettani tulajdonságban a növényekben, állatokban és emberekben.
superdominance - a tulajdonság erősebb megnyilvánulása heterozigóta egyénben (Aa), mint bármelyik homozigóta (AA és aa). Feltételezzük, hogy ez a jelenség heterózis alapját képezi (lásd a 3.7. Bekezdést).
Kodaminirvanie- mindkét allél részvétele a vonás meghatározásában heterozigóta egyénben. A kódolás élénk és jól tanulmányozott példája lehet a negyedik vércsoport öröklése emberben (AB csoport).
Az ebbe a csoportba tartozó emberek vörösvértesteiben kétféle antigén van: antigén A (az egyik kromoszómában jelen lévő gén által / meghatározva) és az antigén az (a / a gén határozza meg, amely egy másik homológ kromoszómán található). Csak ebben az esetben mindkét allél kinyilvánítja hatását - 1 A (A a homozigóta állapot a II. vércsoportot, A) csoportot és én B (homozigóta állapotban a III vércsoportot, B csoportot szabályozza). allél 1 A és én B dolgozzon heterozigótában, mintha egymástól függetlenül működnének.
Öröklési példa csoportok vér szemlélteti többszörös allizmus: gén / három különböző allél képviselhető, de vannak olyan gének, amelyekben több tucat allél van. Egy gén összes allélját nevezik több allél sorozat, amelyekből minden diploid szervezet rendelkezik bármilyen két alléllel (és csak). Ezen allélok között az alléli kölcsönhatások felsorolt \u200b\u200bösszes változata lehetséges.
A több allélizmus jelensége általános a természetben. Számos több allél sorozat ismert, amelyek meghatározzák a kompatibilitást a gombákban történő megtermékenyítés során, a beporzás a vetőmagokban, meghatározzák az állati szőr színét stb.
Nem allél génkölcsönhatások: A nem allél génkölcsönhatásokat számos növényben és állatban leírták. A fenotípus szerint szokatlan felosztású diheterozigóták utódjaiban megjelennek: 9: 3: 4; 9: 6: 1; 13: 3; 12: 3: 1; 15: 1, azaz az általános Mendel-képlet módosítása 9: 3: 3: 1. Két, három vagy több nem allél gén kölcsönhatásának esetei ismertek. Közül a következő fő típusokat lehet megkülönböztetni: komplementeritás, episztázis és polimerizáció.
kiegészítő, vagy további, ezt a nem alléli domináns gének kölcsönhatását nevezik, amelynek eredményeként olyan jel jelenik meg, amely hiányzik mindkét szülőben. Például, amikor az édes borsó két fajtáját keresztezik fehér virágokkal, lila virágokkal rendelkező utódok jelennek meg. Ha megjelöli egy fajta genotípusát AAbb, és a másik - aABB, az
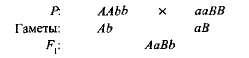
Első generációs hibrid két domináns génnel (A és in) biokémiai alapot kapott a lila pigment antocianin előállításához, bár egyetlen gén sem volt és egyik B gén sem biztosítja e pigment szintézisét. Az antocianin szintézis a nem szekvenciális biokémiai reakciók komplex lánca, amelyet több nem-allél gén irányít, és csak legalább két domináns génnel (A-B) lila szín alakul ki. Más esetekben (aaV- és A-bb) a növény virágai fehérek (a „-” jel a genotípus képletében azt jelzi, hogy mind a domináns, mind a recesszív allélok elfoglalhatják ezt a helyet).
Az édes borsó növényeinek önbeporzása F 1 -ban F 2 megoszlást lila és fehér virágos formákba 9: 7 közeli arányban figyeltünk meg. Lila virágokat találtak Romániában 9/1 6 növény, fehér - 7/16-kor. A Pennet-rács világosan megmutatja ennek a jelenségnek az okát (3.6 ábra).
epistasis - Ez egy olyan génkölcsönhatás, amelyben az egyik gén allélei elnyomják egy másik gén allélpárának megnyilvánulását. gének más gének gátló hatásait nevezzük episztatikus, inhibitorok vagy csökkentők. Az elnyomott gént nevezzük hypostatic.
A fenotípus és egyes osztályok számának és arányának változása szerint a dihibrid hasítás során F 2 vegye figyelembe az episztatikus interakciók több típusát: domináns episztázis (A\u003e B vagy B\u003e A) 12: 3: 1 hasítással; recesszív episztázis (a\u003e B vagy b \u003e A), amelyet 9: 3: 4 osztással stb. fejeznek ki
polymerism amely abban a tényben nyilvánul meg, hogy az egyik jel alul alakul ki befolyása több gén azonos fenotípusos expresszióval. Ezeket a géneket hívják polimer. Ebben az esetben elfogadták a géneknek a tulajdonság kialakulására gyakorolt \u200b\u200begyedi hatásának elvét. Például, amikor egy Pásztorzsák-növényeket kereszteznek háromszögletes és ovális gyümölcsökkel (hüvelyek) az F 1-ben, háromszög alakú gyümölcsök alakulnak ki. Az önbeporzással F 2 megfigyeltük, hogy háromszögletes és ovális hüvelyekkel növényekre osztódnak 15: 1 arányban. Ennek oka az, hogy két gén működik egyedileg. Ezekben az esetekben azonos módon vannak jelölve A 1 és A 2 .
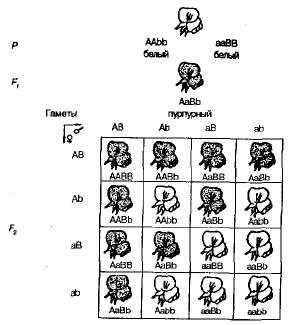
Ábra. 3.6 . A virágok színezésének öröklése édes borsóban
Aztán az összes genotípus (A 1 ,-A 2 ,-, A 1 s 2 és 2 , a 1 egy 1 A 2 -) azonos fenotípusú lesz - háromszög alakú hüvelyek, és csak növények és 1 és 1 és 2 egy 2 különböznek - formálnak ovális hüvelyeket. Ez a helyzet nem kumulatív polimer.
A polimer gének típusonként működhetnek kumulatív polimer. Minél több ilyen gén van a test genotípusában, annál erősebb a tulajdonság megnyilvánulása, azaz a géndózis növekedésével (A 1 A 2 A 3 stb.) tevékenységét összegzik vagy halmozzák. Például a búzaszem endospermiumjának színintenzitása arányos a különféle gének domináns alléljainak számával a trihybrid keresztekben. A legszínezettebbek a gabonafélék voltak A 1 A 1 A 2 A 2 A 3 , A 3 és gabona a 1 és 1 és 2 egy 2 és 3 és 3 nem volt pigment.
A kumulatív polimer típusa szerint sok tulajdonság örökölhető: tejtermelés, tojástermelés, testtömeg és a haszonállatok egyéb jellemzői; a személy fizikai erejének, egészségének és mentális képességeinek számos fontos paramétere; tüske hossza gabonafélékben; cukorrépa a cukorrépa vagy a lipidek gyökerében napraforgómag és t. d.
Így számos megfigyelés azt mutatja, hogy a legtöbb tulajdonság megnyilvánulása az egymással kölcsönhatásba lépő gének komplexumának és a környezeti feltételeknek az egyes jellemzők kialakulására gyakorolt \u200b\u200bhatásának eredménye.
Gén kölcsönhatás
A gének és a tulajdonságok közötti kapcsolat meglehetősen összetett. A testben egy gén nem mindig határozza meg csak egy tulajdonságot, és fordítva: egy tulajdonságot csak egy tulajdonság határoz meg. Gyakrabban egy gén hozzájárulhat több tulajdonság megnyilvánulásához egyszerre, és fordítva. A szervezet genotípusa nem tekinthető független gének egyszerű összegének, amelyek mindegyike egymástól függetlenül működik. Az egyik vagy másik tulajdonság fenotípusos megnyilvánulása sok gén kölcsönhatásának eredménye.
Több génhatás (pleiotropia) - egy génnek a több karakter kialakulására gyakorolt \u200b\u200bhatása.
Például az emberekben a haj vörös színét meghatározó gén világosabbá teszi a bőrt és a szeplők megjelenését.
A morfológiai tulajdonságokat meghatározó gének néha befolyásolják a fiziológiás funkciókat, csökkentve az életerőt és a termékenységet, vagy halálos kimenetelűek. Tehát a gomba, amely a nyér kék színét okozza, csökkenti a termékenységet. A homokigóta állapotban lévő asztrádán juhban a szürke szín domináns génje részletes, mivel ezekben a bárányokban a gyomor alulfejlett és elpusztulnak, amikor a fűtáplálásra váltnak.
A gének komplementer kölcsönhatása. Egy tulajdonság kialakulását több gén is befolyásolhatja. Több nem allél gén kölcsönhatását, amely egy tulajdonság kialakulásához vezet, komplementernek nevezik. Például a csirkéknek négy formája van a fésűnek, ezek bármelyikének megnyilvánulása két nem-allél gén kölcsönhatásával jár. A rózsaszínű héjat az egyik allél domináns génje okozza, a borsószerű héját a másik allél domináns génje okozza. A hibridekben két domináns nem allél gén jelenlétében diószerű héj alakul ki, és minden domináns gén hiányában, azaz két nem alléles gén recesszív homozigótajában egyszerű gerinc alakul ki.
A gének interakciójának eredménye a szőr színezése kutyákban, egerekben, lovakban, a sütőtök alakja, az édes borsóvirág színe.
A polimer olyan nem allél gének interakciója, amikor a tulajdonság fejlõdésének mértéke a domináns gének teljes számától függ. Ezen elv szerint a zab- és búzaszem színét, valamint az ember bőrének színét öröklik. Például, a felekben két pár nem-alléles génben 4 domináns gén található, míg a fehér bőrű emberekben - egy sem - az összes gén recesszív. A domináns és recesszív gének különféle számainak kombinációja olyan mulatto kialakulásához vezet, amelynek bőrszín-intenzitása különbözik: sötéttől világosig.
A gén-interakció két fő csoportja van: az allél gének közötti kölcsönhatás és a nem allél gének közötti kölcsönhatás. Meg kell érteni azonban, hogy ez nem a gének fizikai kölcsönhatása, hanem az elsődleges és a másodlagos termékek kölcsönhatása, amely meghatározza az egyik vagy a másik tulajdonságot. A citoplazmában kölcsönhatás lép fel a fehérjék között - enzimek között, amelyek szintézisét a gének határozzák meg, vagy az anyagok között, amelyek ezen enzimek hatására képződnek.
A következő típusú interakciók lehetséges:
1) egy specifikus tulajdonság kialakulásához két enzim kölcsönhatása szükséges, amelyek szintézisét két nem allél gén határozza meg;
2) az egyik gén részvételével szintetizált enzim teljesen elnyomja vagy inaktiválja annak az enzimnek a hatását, amelyet egy másik nem allél gén alkotott;
3) két enzim, amelyek képződését két nem allél gén szabályozza, amelyek befolyásolják egy tulajdonságot vagy egy folyamatot úgy, hogy együttes hatása a tulajdonság megnyilvánulásának megjelenéséhez és fokozódásához vezet.
Allerikus gén kölcsönhatás
Azokat a géneket, amelyek azonos (homológ) lókuszt foglalnak el a homológ kromoszómákon, allélnak nevezik. Mindegyik szervezetnek két allélgénje van.
Az allél gének közötti interakció ilyen formái ismertek: teljes dominancia, hiányos dominancia, kódolás és túlhatás.
Az interakció fő formája a teljes dominancia, amelyet először G. Mendel írt le. Lényege abban rejlik, hogy egy heterozigóta szervezetben az egyik allél megnyilvánulása uralja a másik megjelenését. Az 1: 2: 1 genotípusos hasítás teljes dominanciájával nem egybeesik a 3: 1 fenotípusos hasításával. Az orvosi gyakorlatban kétezer monogén örökletes betegség közül csaknem a felében van a patológiás gének domináns megnyilvánulása a normáléhoz képest. Heterozigótákban a patológiás allél a legtöbb esetben a betegség jeleivel (domináns fenotípus) nyilvánul meg.
A hiányos dominancia egy olyan interakció olyan formája, amelyben a heterozigóta szervezetben (Aa) az domináns (A) gén nem szünteti meg teljesen az (a) recesszív gént, amelynek eredményeként a közbenső karakterek között köztitermék jelenik meg. Ebben az esetben a genotípus és a fenotípus megoszlása \u200b\u200begybeesik, és 1: 2: 1 arányú
Ha heterozigóta organizmusokban kódolnak, akkor az allélek mindegyikében egy-egy termék képződését okozza, amely attól függ, vagyis mindkét allél termékei megtalálhatók. Ennek a megnyilvánulásnak a klasszikus példája a vércsoport-rendszer, különösen az ABO-rendszer, amikor az emberi vörösvérsejtek antigént hordoznak a felületen, amelyet mindkét allél szabályoz. A megnyilvánulásnak ezt a formáját kódolásnak nevezik.
Túlsúly: amikor egy domináns gén heterozigóta állapotban kifejezettebb, mint egy homozigóta állapotban. Tehát Drosophila-ban az AA genotípusú normál élettartam; Aa - az élet meghosszabbított trivatizmusa; aa - halál.
Többszörös aleizmus
Mindegyik szervezetnek csak két allélgénje van. A természetben azonban az allélok száma kettőnél is több lehet, ha egyes lókuszok különböző állapotokban lehetnek. Ilyen esetekben több allélról vagy több allelomorfizmusról beszélnek.
Több allélt azonos betű jelöl, különféle indexekkel, például: A, A1, A3 ... Az allél gének a homológ kromoszómák azonos részeiben vannak lokalizálva. Mivel a kariotípusban mindig két homológ kromoszóma van jelen, akkor több allél esetén minden szervezetnek egyszerre csak két azonos vagy eltérő allélje lehet. Csak egyikük lép be a reproduktív sejtbe (a homológ kromoszómák különbségével együtt). Több allél esetén az összes allél jellemző tulajdonsága ugyanazon a tulajdonságon. A különbség közöttük csak a megjelölés fejlettségi szintje mutatkozik.
A második tulajdonság az, hogy a szomatikus sejtekben vagy a diploid organizmusok sejtjeiben több allél közül kettő legfeljebb két található, mivel ezek a kromoszóma azonos lokuszában vannak.
A több allélban rejlő másik jellemző. A dominancia jellege alapján az allelomorf karakterek sorrendben vannak elrendezve: a normál, változatlan karakter uralja a többieket, a sorozat második génje az elsőhez képest recesszív, de a következőket uralja stb. A több allél emberben történő megnyilvánulásának egy példája az ABO-rendszer vércsoportja.
A többszörös aleizmus nagy biológiai és gyakorlati jelentőséggel bír, mivel fokozza a kombinatív variabilitást, különös tekintettel a genotípusra.
Nem egy gén kölcsönhatás
Sok olyan eset van, amikor egy tulajdonságot vagy tulajdonságot két vagy több elválaszthatatlan gén határozza meg, amelyek kölcsönhatásba lépnek egymással. Bár itt az interakció feltételes, mivel nem a gének, hanem az általuk ellenőrzött termékek kölcsönhatásba lépnek. Ebben az esetben eltérés mutatkozik a felosztás mendelivi törvényeitől.
A géninterakciónak négy fő típusa van: komplementaritás, episztázis, polimerizáció és módosító hatás (pleiotropia).
A komplementaritás a nem allél gének interakciójának egy típusa, amikor az egyik domináns gén kiegészíti egy másik nem alléli domináns gén hatását, és együttesen meghatároznak egy új tulajdonságot, amely a szülőkben hiányzik. Sőt, a megfelelő tulajdonság csak mindkét nem allél gén jelenlétében alakul ki. Például az egerek kabátjának kénszínét két gén (A és B) szabályozza. Az A gén határozza meg a pigment szintézisét, de mind a homozigóták (AA), mind a heterozigóták (Aa) albínók. Egy másik B gén pigmentek halmozódását biztosítja, főleg a haj alján és végén. A digeterozigóták (AabB x AabB) keresztezése a hibridek 9: 3: 4 arányú hasadásához vezet. A komplementer kölcsönhatások számaránya akár 9: 7 lehet; 9: 6: 1 (a Mendeliv felosztás módosítása).
A humán komplementer génkölcsönhatásokra példa a védőfehérje - interferon szintézise. A testben történő kialakulása a nem-allél gének komplementer kölcsönhatásával jár, amelyek különböző kromoszómákon helyezkednek el.
Az episztazis egy nem allél gének kölcsönhatása, amelyben az egyik gén gátolja egy másik nem allél gén hatását. Mind a domináns, mind a recesszív gének (A\u003e B, a\u003e B, B\u003e A, B\u003e A) gátlást okozhatnak, és ettől függően az episztatist dominánsnak és recesszívnek tekintik. A gátló gént inhibitornak vagy szuppresszornak nevezzük. Az inhibitorgének általában nem határozzák meg egy adott tulajdonság kialakulását, hanem csak gátolják egy másik gén hatását.
Egy gént, amelynek hatása elnyomódik, hiposztatikusnak nevezzük. Az episztatikus génkölcsönhatások mellett az F2 fenotípushasadása 13: 3; 12: 3: 1 vagy 9: 3: 4 és mások: A tök gyümölcsének színét és a lovak színét az ilyen típusú kölcsönhatás határozza meg.
 Gyere vagy menj? Mi a különbség
Gyere vagy menj? Mi a különbség A születési órát a Ba Zi kártyában határozzuk meg - "Az élet jó" - az élet pozitív gondolkodása Feng Shui szerint
A születési órát a Ba Zi kártyában határozzuk meg - "Az élet jó" - az élet pozitív gondolkodása Feng Shui szerint Sirius jelentősen befolyásolta és továbbra is befolyásolja a föld - föld történeteket
Sirius jelentősen befolyásolta és továbbra is befolyásolja a föld - föld történeteket