Дали женският пол винаги се определя от комбинация от xx. Подова биология
Генетика на пода
Полът се характеризира с комплекс от признаци, определени от гените, разположени в хромозомите. В клетките на човешкото тяло, хромозомите са двойни диплоидни комплекти. При видове с двудомни индивиди, хромозомният комплекс на мъжките и женските не е еднакъв и се различава с една двойка хромозоми (полови хромозоми). Идентичните хромозоми от тази двойка се наричат X (X) -хромозома, несдвоен, отсъстващ от другия пол-Y (игра) -хромозома; останалите, за които няма разлики, са автозоми (А).
Женските клетки съдържат два идентични полови хромозоми, обозначени като ХХ, като при мъжете те са представени от две несвързани хромозоми Х и Y. Така че, наборът от хромозоми на мъж и жена се различава само в една хромозома: хромозомният комплект на жената съдържа 44 автозоми + ХХ, мъжете - 44 автозоми + XY.
При разделянето и узряването на зародишните клетки при хората се образуват гамети с хаплоиден брой хромозоми: яйцата по правило съдържат 22 + Х хромозоми. Така при жените се образува само един вид гамета (гамети с Х-хромозомата). При мъжките гамети съдържат 22 + X или 22 + Y хромозоми и се образуват два вида гамети (гамета с Х хромозома и гамета с Y хромозома). Ако по време на оплождането сперматозоида с Х-хромозома влезе в яйцеклетката, се образува женски зародиш, а с Y хромозомата е мъжки.
Следователно определянето на пола на човек зависи от наличието в мъжките зародишни клетки - сперматозоиди, оплождащи яйцето, Х- или Y-хромозомите.
Има четири основни вида определяне на хромозомния пол:
1. Мъжкият пол е хетерогаметен; 50% от гаметите носят Х-, 50% -У - хромозома, например човек, бозайници, диплопи, бръмбари, бъгове (Слайд 4).
2. Мъжкият пол е хетерогаметен; 50% от гаметите са Х-, 50% нямат полова хромозома, например скакалци, кенгуру (Слайд 7).
3. Женският пол е хетерогаметен; 50% от гаметите носят Х-, 50% гамета –Y– хромозома, например птици, влечуги, опашки земноводни, копринени буби (Слайд 7).
4. Женският пол е хетерогаметен; 50% от гаметите носят Х-, 50% нямат полова хромозома, например, мол.
Наследяването на признаци, чиито гени са локализирани в половите хромозоми, се нарича наследяване, свързан с пода.
26. Генотипът като цялостна система. Взаимодействието на гените, множественото действие на гените.
Генотипът като холистична система
Свойства на гените. Въз основа на търси с примери за наследяване на черти при моно- и ди-хибридни кръстове може да се създаде впечатлението, че генотипът на организма се състои от сумата от отделни, независимо действащи гени, всеки от които определя развитието на собствената си черта или свойство. Такава идея за пряка и недвусмислена връзка на гена с черта най-често не е вярна. В действителност, има огромен брой признаци и свойства на живите организми, които се определят от две или повече двойки гени, и обратното, един ген често контролира много признаци. В допълнение, ефектът на един ген може да бъде променен от близостта на други гени и условия на околната среда. Така в онтогенезата не действат отделни гени, а целият генотип като цялостна система със сложни връзки и взаимодействия между неговите компоненти. Тази система е динамична: появата на нови алели или гени в резултат на мутации, образуването на нови хромозоми и дори новите геноми води до забележима промяна в генотипа с течение на времето.
Естеството на проявата на генното действие в състава на генотипа като система може да варира в различни ситуации и под влиянието различни фактори. Това може лесно да се види, ако разгледаме свойствата на гените и характеристиките на тяхното проявление в знаците:
Генът е дискретен в действието си, т.е. той е изолиран в своята активност от други гени.
Генът е специфичен в своята проява, т.е. отговаря за строго определена характеристика или свойство на организма.
Генът може да действа постепенно, т.е. да повишава степента на проявление на черта с увеличаване на броя на доминантните алели (генна доза).
Един единствен ген може да повлияе на развитието на различни черти - това е множествен, или плейотропен, ефект на ген.
Различните гени могат да имат същия ефект върху развитието на една и съща черта (често количествени характеристики) - те са множество гени или полигени.
Генът може да взаимодейства с други гени, което води до появата на нови признаци. Такова взаимодействие се извършва непряко - чрез продуктите на техните реакции, синтезирани под техен контрол.
Действието на гена може да бъде променено чрез промяна на местоположението му на хромозомата (позиционен ефект) или чрез влиянието на различни фактори на околната среда.
Взаимодействия на алелни гени. Феноменът, когато няколко гена (алели) са отговорни за една черта, се нарича генно взаимодействие. Ако това са алели на един и същи ген, тогава се наричат такива взаимодействия алелен, и в случай на алели от различни гени - неалелни.
Разграничават се следните основни типове алелни взаимодействия: доминантност, непълна доминантност, свръхдоминантност и кодоминантност.
господство - вида на взаимодействие на два алела на един ген, когато един от тях напълно елиминира проявлението на другия. Такъв феномен е възможен при следните условия: 1) доминиращият алел в хетерозиготното състояние осигурява синтез на продукти, достатъчни за проявяване на черта с същото качество, както в състоянието на доминантната хомозигота в родителската форма; 2) рецесивният алел е напълно неактивен, или продуктите от неговата активност не взаимодействат с продуктите на активността на доминиращия алел.
Примери за това взаимодействие на алелните гени са доминирането на лилавото цвят цветя на грах над бяла, гладка форма на семена над набръчкана, тъмна коса над светлина кафяви очи синьо в хората и т.н.
Непълно доминиране или междинен характер на наследството наблюдавано в случая, когато фенотипът на хибрида (хетерозиготи) се различава от фенотипа на двете родителски хомозиготи, т.е., експресията на признака е междинна, с повече или по-малко отклонение към един или друг родител. Механизмът на това явление е, че рецесивният алел е неактивен, а степента на активност на доминиращия алел е недостатъчна, за да осигури желаното ниво на проявление на доминантната черта.
Пример за непълно господство е наследството. цвят цветя в растенията от нощната красота (фиг. 3.5). Както може да се види от диаграмата, хомозиготни растения имат или червено (АА) или бяло (Аа) цветя и хетерозиготни (Аа) - розово. При пресичане на растения с червени цветя и растения с бели цветя във F 1, всички растения имат розови цветя, т.е. има междинен характер на наследството. При преминаване на хибриди с розови цветя F 2 има съвпадение на разцепването с фенотип и генотип, тъй като доминиращата хомозигота (АА) различен от хетерозиготите (Аа). Така че, в този пример с нощните козметични растения, разделяйки се F 2 Цветът на цветята обикновено е следният - 1 червено (АА): 2 розов (Аа): 1 бяло (Аа).
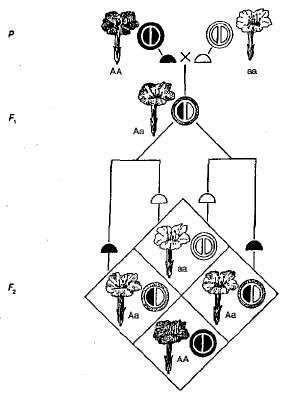
Фиг. 3.5. Наследяване на цвят на цветя в случай на непълно превъзходство на нощната красота.
Непълното доминиране е широко разпространено. Наблюдава се в наследяването на къдрава коса при хората, цвета на едрия рогат добитък, цвета на оперението при пилетата и много други морфологични и физиологични признаци в растенията, животните и хората.
superdominance - по-силно проявление на чертата в хетерозиготен индивид (Аа), от всеки хомозигот (АА и аа). Предполага се, че това явление е в основата на хетерозис (виж § 3.7).
Kodaminirvanie- участието на двата алела при определянето на признака в хетерозиготен индивид. Ярък и добре проучен пример за кодоминантност е наследяването на кръвна група IV при хора (група АВ).
Червените кръвни клетки на хората от тази група имат два вида антигени: антиген А (определен от генома / в един от хромозомите) и антиген Най- (определен от генома / а, локализиран върху друга хомоложна хромозома). Само в този случай и двата алела - 1 А (В хомозиготно състояние контролира кръвна група II, група А) и аз B (в хомозиготно състояние контролира III кръвна група, група В). алели 1 А и аз B работа в хетерозигота, сякаш независимо един от друг.
Пример за наследяване групи кръв илюстрира и проявява множествен алел: ген / може да бъде представен от три различни алела и има гени, които имат десетки алели. Всички алели на един и същи ген се наричат серия от множество алели, от които всеки диплоиден организъм може да има две алели (и само). Между тези алели са възможни всички изброени варианти на алелни взаимодействия.
Феноменът на множествения алел е често срещан в природата. Съществуват обширни серии от многобройни алели, които определят вида на съвместимост по време на оплождането при гъби, опрашване в семената, определяне на цвета на животинския косъм и др.
Взаимодействия на неалелни гени В много растения и животни са описани взаимодействия с не-алелни гени. Те водят до появата в поколението на дигерозитовете на необичайно разцепване по фенотип: 9: 3: 4; 9: 6: 1; 13: 3; 12: 3: 1; 15: 1, т.е. модификации на общата формула на Мендел 9: 3: 3: 1. Има случаи на взаимодействие на два, три и повече неалелни гени. Сред тях са следните основни видове: комплементарност, епистаза и полимери.
комплементарна, или допълнително, Това взаимодействие се нарича неалелни доминантни гени, в резултат на което се появява черта, която липсва при двамата родители. Например, когато две разновидности на сладък грах са кръстосани с бели цветя, потомството се появява с лилави цветя. Ако обозначите генотипа на един сорт AABB, и другото aABB, на
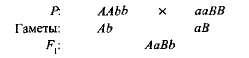
Хибрид от първо поколение с два доминантни гена (А и В) получил биохимична основа за производството на антоцианин лилав пигмент, докато или един по един ген И, Нито един от двата гена В осигурява синтеза на този пигмент. Синтезът на антоцианин е сложна верига от последователни биохимични реакции, контролирани от няколко неалелни гени и само ако има поне два доминантни гена (А-В) се развива лилав цвят. В други случаи (aaV- и А-бб) цветята на растението са бели (знакът "-" в формулата на генотипа означава, че това място може да бъде заето както от господстващия, така и от рецесивния алел).
При самоопрашващи се растения от сладък грах F 1 в F 2 разделянето на пурпурни и бялоцветни форми се наблюдава в съотношение, близко до 9: 7. В него са намерени пурпурни цветя 9/1 6 растения, бяло - на 7/16. Решетката на Pennet ясно показва причината за това явление (фиг. 3.6).
epistasis - това е вид генно взаимодействие, при което алелите на един ген потискат проявата на алелна двойка от друг ген. гени инхибират действието на други гени епистатични инхибитори или заглушители. Потисканият ген се нарича ипостасното.
Според промяната в броя и съотношението на фенотипа и цикличните класове по време на разделянето на дихибрид F 2 разгледайте няколко вида епистатични взаимодействия: доминираща епистаза (A\u003e B или B\u003e A) с разцепване 12: 3: 1; рецесивна епистаза (а\u003e Б или b \u003e А), което се изразява в разделяне 9: 3: 4 и т.н.
polymerism проявява се във факта, че под една форма се формира един знак влиянието няколко гена със същата фенотипна експресия. Тези гени се наричат полимер. В този случай е възприет принципът на недвусмислено действие на гените върху развитието на признака. Например, при пресичането на растенията с овчарски торбички с триъгълни и овални плодове (шушулки) във F 1 се образуват растения с плодове с триъгълна форма. С тяхното самоопрашване F 2 наблюдава се разцепване на растения с триъгълни и овални шушулки в съотношение 15: 1. Това се обяснява с факта, че съществуват два гена, които действат еднозначно. В тези случаи те се означават идентично. А 1 и А 2 .
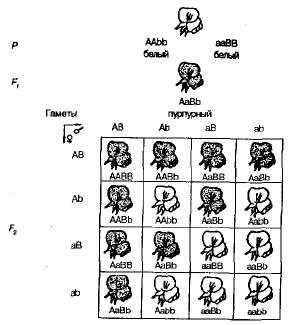
Фиг. 3.6 , Наследяване на цвят цвете в сладък грах
След това всички генотипове (А 1 ,-А 2 ,-, А 1 ите 2 и 2 , а 1 а 1 А 2 -) ще имат същия фенотип - триъгълни шушулки и само растения и 1 и 1 и 2 а 2 ще бъде различно - да образуват овални шушулки. Такъв е случаят некумулативни полимери.
Полимерните гени могат да действат по вид кумулативни полимери. Колкото по-сходни са гените в генотипа на организма, толкова по-силно е проявлението на тази черта, т.е. с увеличаване на дозата на гена (А 1 А 2 А 3 и т.н.) действието му е обобщено или натрупано. Например, интензивността на цвета на ендосперма на зърната на пшеницата е пропорционална на броя на доминиращите алели на различни гени при трихибридно преминаване. Най-оцветените зърна бяха А 1 А 1 А 2 А 2 А 3 , А 3 и зърно a 1 и 1 и 2 а 2 и 3 и 3 нямаше пигмент.
Според вида на кумулативния полимер се наследяват много признаци: млечност, производство на яйца, тегло и други признаци на селскостопански животни; много важни параметри на физическата сила, здраве и умствени способности на човека; дължината на зърното на ухото; съдържание на захар в корените или липидите на захарното цвекло слънчогледови семена и така нататък
По този начин многобройните наблюдения показват, че проявлението на повечето от признаците е резултат от влиянието на комплекс от взаимодействащи гени и условия на околната среда върху формирането на всяка специфична черта.
Генно взаимодействие
Връзката между гените и чертите е доста сложна. В един организъм не винаги един ген определя само една черта и, обратно, една черта се определя само от един ген. По-често един ген може да допринесе за проявлението на няколко признака наведнъж и обратно. Генотипът на организма не може да се разглежда като проста сума от независими гени, всяка от които функционира извън допира с другите. Фенотипните прояви на черта са резултат от взаимодействието на много гени.
Множествено генно действие (плейотропия) - процесите на влияние на един ген върху образуването на няколко символа.
Например, при хората, ген, който определя червения цвят на косата, причинява по-светла кожа и лунички.
Понякога гените, които определят морфологичните характеристики, засягат физиологичните функции, намаляват жизнеността и плодовитостта или са фатални. По този начин ген, който причинява син цвят в норка, намалява плодовитостта му. Доминиращият ген на сивото оцветяване в аракъл овце в хомозиготно състояние е подробен, тъй като такива агнета имат слабо развит стомах и умират, когато преминат на трева.
Допълнително генно взаимодействие. Няколко гени могат да повлияят на развитието на една черта. Взаимодействието на няколко неалелни гени, водещи до развитието на единична черта, се нарича допълващо. Например, пилетата имат четири форми на гребена, проявлението на който и да е от тях е свързано с взаимодействието на две двойки неалелни гени. Розовият гребен се дължи на действието на господстващия ген на един алел, гребеновидният гребен - доминиращият ген на другия алел. При хибридите, в присъствието на два доминантни неалелни гена, се образува ореховиден гребен и при липса на всички доминантни гени, т.е. в рецесивни хомозиготи за два неалелни гена се образува обикновен гребен.
Резултатът от взаимодействието на гените е оцветяването на вълна при кучета, мишки, коне, формата на тиква, цветът на цветята на сладък грах.
Полимерът е взаимодействие на неалелни гени, когато степента на развитие на черта зависи от общия брой на доминантните гени. Според този принцип, оцветяването на зърната от овес и пшеница и цветът на кожата на човек се наследяват. Например, в черните има 4 доминантни гена в два чифта неалелни гени и никой при хора с бяла кожа, всички гени не са рецесивни. Комбинации от различни количества доминиращи и рецесивни гени води до образуването на мулати с различна интензивност на цвета на кожата: от тъмни към светли.
Има две основни групи на генно взаимодействие: взаимодействието между алелните гени и взаимодействието между не-алелните гени. Въпреки това, трябва да се разбере, че това не е физическото взаимодействие на самите гени, а взаимодействието на първични и вторични продукти, които ще определят една или друга черта. В цитоплазмата се осъществява взаимодействие между протеини - ензими, чийто синтез се определя от гени, или между вещества, които се образуват под влиянието на тези ензими.
Възможни са следните видове взаимодействие:
1) за образуването на специфична черта е необходимо взаимодействие на два ензима, синтезът на който се определя от два не-алелни гена;
2) ензимът, който е синтезиран с участието на един ген, напълно потиска или инактивира действието на ензима, който се формира от друг неалелен ген;
3) два ензима, образуването на които се контролира от два не-алелни гена, които засягат една черта или един процес, така че съвместното им действие води до появата и усилването на проявата на чертата.
Взаимодействие с алелен ген
Гените, които заемат идентични (хомоложни) локуси в хомоложни хромозоми, се наричат алелни. Всеки организъм има два алелни гена.
Такива форми на взаимодействие между алелни гени са известни: пълно господство, непълно господство, кодоминантност и наддоминантност.
Основната форма на взаимодействие е пълно господство, което за първи път е описано от G. Mendel. Нейната същност е в това, че в хетерозиготен организъм проявлението на един от алелите доминира над проявлението на другия. При пълното доминиране на разцепването по генотип 1: 2: 1 не съвпада с разделянето по фенотип - 3: 1. В медицинската практика, от две хиляди моногенни наследствени заболявания почти наполовина има доминиращо проявление на патологични гени над нормалните. При хетерозиготите патологичният алел се проявява в повечето случаи с признаци на болестта (доминантния фенотип).
Непълното доминиране е форма на взаимодействие, при която доминиращият ген (А) в хетерозиготен организъм (Аа) не потиска напълно рецесивния ген (а), в резултат на което се появява междинен знак между родителските знаци. Тук разделянето на генотипа и фенотипа е същото и е 1: 2: 1
Когато кодоминант в хетерозиготни организми, всеки от алелните гени причинява образуването на продукт, зависим от него, т.е. продуктите на двата алела се оказват. Класически пример за такава проява е системата на кръвните групи, по-специално системата ABO, когато човешките еритроцити носят антигени на повърхността, които се контролират от двата алела. Тази форма на проявление се нарича codominance.
Супердоминантност - когато доминантният ген е по-изразен в хетерозиготно състояние, отколкото в хомозиготно състояние. Така, в Drosophila, с генотип АА, нормалната продължителност на живота; Аа - удължен живот на триватистите; аа - смъртоносен.
Многобройни алеизми
Всеки организъм има само два алелни гена. Въпреки това, често в природата, броят на алелите може да бъде повече от две, ако някои локус може да бъде в различни състояния. В такива случаи се казват множество алели или множество алеломорфизми.
Множество алели са обозначени с една и съща буква с различни индекси, например: А, А1, А3 ... Анелните гени са разположени в същите области на хомоложни хромозоми. Тъй като винаги има два хомоложни хромозоми в кариотипа, дори и с множество алели, всеки организъм може да има едновременно само два еднакви или различни алела. Само една от тях влиза в репродуктивната клетка (заедно с разликата в хомоложните хромозоми). За множество алели, характерният ефект на всички алели върху една и съща черта. Разликата между тях е само в степента на развитие на чертата.
Втората особеност е, че соматичните клетки или клетките от диплоидни организми съдържат максимум две алели от няколко, тъй като те се намират в един и същ хромозомен локус.
Друга особеност е присъща на множество алели. По характер на господство, алоломорфните знаци се поставят в последователен ред: по-често нормалната, непроменена черта доминира над другите, вторият ген от серията е рецесивен по отношение на първия, но доминира следното и т.н. Пример за проявление на множествени алели в лицето са кръвните групи на системата ABO.
Множественото алеизъм има важно биологично и практическо значение, тъй като увеличава комбинираната вариабилност, особено генотипната.
Взаимодействие на не-алелни гени
Има много случаи, при които черта или свойства се определят от два или повече неотделими гена, които взаимодействат помежду си. Въпреки че тук взаимодействието е условно, защото не са взаимодействащи гените, а продуктите, които те контролират. В този случай, има отклонение от моделите на Менделско разделяне.
Съществуват четири основни типа взаимодействия на гените: комплементарност, епистаза, полимери и модифициращо действие (плейотропия).
Допълняемостта е такъв тип взаимодействие на неалелни гени, когато един господстващ ген допълва действието на друг неалелен господстващ ген и заедно определят нова черта, която липсва в родителите. Освен това, съответната черта се развива само в присъствието на двата не-алелни гена. Например, сярата е цветът на вълната при мишки, контролирани от два гена (А и В). Ген А определя пигментния синтез, но хомозиготите (АА) и хетерозиготите (Аа) са албинози. Друг ген В осигурява пигментни клъстери главно в основата и по върховете на косата. Кръстосването на дихетерозиготи (AaBb x AaBb) води до разделяне на хибридите в съотношение 9: 3: 4. Числените съотношения за допълнителни взаимодействия могат да бъдат 9: 7; 9: 6: 1 (модификация на разделянето на Менделеев).
Пример за допълнителното взаимодействие на гените при хората може да бъде синтеза на защитен протеин, интерферон. Образуването му в тялото е свързано с допълващото взаимодействие на два не-алелни гена, разположени в различни хромозоми.
Епистазата е взаимодействие на неалелни гени, в които един ген потиска действието на друг неалелен ген. И доминантните, и рецесивните гени могат да причинят депресия (A\u003e B, A\u003e B, B\u003e A, B\u003e A) и в зависимост от това епистазата е доминираща и рецесивна. Преобладаващият ген се нарича инхибитор или супресор. Инхибиторните гени обикновено не определят развитието на определена черта, а само инхибират действието на друг ген.
Генът, чийто ефект е потиснат, се нарича хипостатичен. При епистатично генно взаимодействие, разцепването на фенотипа във F2 е 13: 3; 12: 3: 1 или 9: 3: 4 и др. Цветът на плодовете от тиква, цветът на конете се определя от този вид взаимодействие.
Пол - това е набор от морфологични, физиологични, биохимични, поведенчески и други признаци на тялото, които причиняват репродукцията.
Признаците, с които се различават индивиди от различни полове, се разделят на _ първични и вторични. Първичните органи са органи, които осигуряват образуването на гамети и оплождането (половите жлези, гениталния тракт, органите). Това са външни и вътрешни генитални органи, които се поставят в ембриогенезата. Вторични - не участват в сексуалното размножаване. Развиват се под влиянието на половите хормони и се появяват в пубертета (при хора след 12-15 години). Това са характеристики на развитието на опорно-двигателния апарат, подкожната мастна тъкан, косата, тембъра на гласа, поведенческите характеристики; при птици, пеене, оперение и др.
Признаците на лица, свързани със секса, могат да бъдат разделени на 3 категории:
1) ограничено до пода
2) контролиран етаж
3) свързан с пода.
Развитието на първия се дължи на гените, разположени в автозомите на двата пола, но се проявяват само в един пол. По този начин, гени за поставяне на яйца присъстват в пилета и петли, но се появяват само при пилета. По същия начин се държат гените на мазнини и млека при говедата. Това явление се наблюдава във връзка с експозицията на съответните полови хормони.
Пример за второто е проявлението на рога в мъжки крави, комолост - в женски. При хора: плешивост при мъже, подагра - 80% при мъже и 12% при жени.
Признаците, чието развитие се контролира от гени на половите хромозоми, се наричат свързани с пола. Такива признаци са около 200. При Х-хромозомата има свързана цветна слепота, хемофилия; Y-хромозома - хипертрихоза, ихтиоза.
ВИДОВЕ ОПРЕДЕЛЯНЕ НА ЕТАЖИТЕ.
1. Прогамен - преди оплождане. Съотношението на половите хромозоми в този случай не играе роля, тъй като диплоидни ооцити. (Някои червеи, ротатори - женски се развиват от големи яйцеклетки, мъжки от малки ооцити).
2. Singamic - генетично определяне на пола по време на оплождането, което зависи от естеството на комбинацията от половите хромозоми или съотношението на половите хромозоми и автозомите.
3. Епигамично - под въздействието на външната среда (червей бонелия).
ХРОМОСОМНА ОПРЕДЕЛЕНИЕ НА ЕТАЖА
┌─────────────────────────┬───────────────────┬──────────────────┐
│ Хромозомни типове │ генотипи │ типове гамета
├ определяне на пола ├─────────┬─────────┼────────┬─────────┤
│ │ │ │ │ │
├─────────────────────────┼─────────┴─────────┼────────┼─────────┤
Hete мъжки хетерогам │
│ Инсекти от щъркели │ │ │ │
Bug (бъгове Protenor, бръмбари, │ │ │ │)
Ukipauki, скакалци) ХО │ ХХ, Х, О │ Х │
Oph Drosophilus Y XY │ XX │ X, Y X │
Br Гръбначни │ │ │
Mam (бозайници, човек) Y XY │ XX, X, Y │ X │
│ │ │ │ │ │
Hete женска хетерогама │ │
S Птици. риба, пеперуди, │ │ │ │
Коп │ │ │ │
. Водни. │ XX Y XY │ X, X, Y │
AndЦена и други безсилни │ │ │ │
Нощ │ XX │ XO │ X │ X, O
└─────────────────────────┴─────────┴─────────┴────────┴─────────┘
Типът на дефиницията на сингама е дефиниция на хромозома гени с генетичен контрол. Отговорен за пола на хромозомата, наречен секс. Нормален мъжки гамета носи или Х или Y хромозома, а всички яйца - Х-хромозомата. В случай на нормално несъответствие на хромозомите по време на мейоза се образуват нормални яйца и сперматозоиди с обичайния набор от хромозоми X и Y. Полът на зиготата се определя от съотношението между гамети XX и XY (хомогаметични и хетерогаметни).
Хромозомната теория на пола на Коренс (1907) е, че полът се определя от комбинацията на половите хромозоми по време на оплождането. Разграничават се следните видове определяне на хромозомния пол: XY, XO, ZW, ZO.
В случай на нарушение на хода на митоза или мейоза, могат да се образуват гинандоморфи. Съдържанието на половите хромозоми в различни клетки на такива индивиди може да бъде различно (мозайка). В Drosophila летят: XX и XO, в човек XX и XY, във връзка с които различни болки в тялото могат да имат съответни признаци на секс. Може да има и други случаи на мозаицизъм: XX / XXX, XY / XXX; XO / XXY и др
Ако половите хромозоми не се разпаднат, може да има 12 възможни комбинации на половите хромозоми в човешките зиготи, което е причина за хромозомни аберации при хората.
│ X. XX │ O
─────┼────────┼─────────┼────────
X │ XX │ XXX O XO
Y Y XY │ XXY. YO
XY Y XXY Y XXXY YO XYO
O O XO │ │
В случай на несъвместимост на половите хромозоми по време на мейоза, при жените се образуват гамети XX и O. и XY и О при мъже. С участието си в оплождането зиготите се формират с необичайна комбинация от полови хромозоми. При хората такива аномалии възникват 1 на 600-700 новородени. Зиготе Йо умира на ранен етап; физически лица XXX, XXY, XO са жизнеспособни и техният пол зависи от наличието или отсъствието на хромозома "Y", която, с произволен брой Х хромозоми, контролира образуването на признаци на мъжкия пол, развитието и стимулира образуването на тестисите. Излишъкът на Х-хромозомите причинява конституционални аномалии и дефекти на интелигентността. Но в природата има индивиди, в които хромозомата „Y” е генетично инертна и няма специално влияние върху определянето на пола.
Бяха открити екземпляри от тип Drosophila от тип CW, които бяха мъжки, но безплодни (1916 г., C. Bridges), а XXY индивидите са нормални фертилни жени.
Балансовата теория на пола (Bridges, 1922). Изучава съотношението между гениталиите и автозомите
При нормални жени с набор от хромозоми 2n, съотношението на автозомни и Х хромозоми е 1: 2n = 2A + 2X (2X: 2A = 1 - нормално женско), 1, 5 - над женското: 2A + 3X (3X: 2A = 1, 5 - безплодни). При мъжете съотношението е 0, 5 2n = 2A + XY (X: 2A = 0, 5). С намаляването на индивида остават мъже 3A + XY (X: 3A = 0, 33 - безплодни) - супермени. Стойността на коефициента между 1 и 0, 5 съответства на фенотипа на междинните индивиди по полова _интекс.: 3A + 2X (2X: 3A = 0, 66 - признаци на двата пола са безплодни).
Следователно, същността на теорията на баланса е, че не само сексуалните хромозоми, но и автозомите участват в определянето на пола. Един хаплоиден набор от автозоми разказва индивидуалните свойства на мъжа. В този случай, етажът се определя от съотношението на броя (баланса) на автозомите и половите хромозоми.
Определянето на пола чрез плоидност се открива и при пчелите. Женските са диплоидни, а мъжките са хаплоидни, защото развиват партеногенетични от неоплодени яйца.
ДИФЕРЕНЦИАЦИЯ НА ЕТАЖА В ОНТОГЕНЕЗА
Началото на половите жлези в ранните ембриони (до 5-та или 6-та седмица) не се различава в различните полове и се състои от външния слой - кора на кората и вътрешния слой - мозъка, не съдържат клетки от зародишна линия. Първичните зародишни клетки се откриват при хора на третата седмица от ембрионалното развитие в ектодермата на жълтъчния сак. След това, под въздействието на хемотаксични сигнали, те мигрират към гонадите. Тази миграция е независима от пола. Основите на половите жлези могат да се развият в яйчниците или тестисите. Диференциацията на гонади се наблюдава на 8-та седмица: на ден 36, тестиса започва да секретира андрогени (тестостерон), което определя развитието на мъжкия пол.
В организмите, които имат половите жлези, се упражнява генетичен контрол върху формирането на половите характеристики.
Гендерната диференциация е програмирана от генетичен набор от полови хромозоми, образувани по време на сливането на женските и мъжките гамети. Генетичният пол на ембриона се определя от половите хромозоми XX или XY.
Посоката на развитие на пола се определя от наличието на Y хромозома. Обикновено Х-хромозомите съдържат репресорен ген (Tfm, тестикуларен феминизационен ген), който предотвратява развитието на мъжки тип. Нормалният алел на гена определя синтеза на протеиновия рецептор за андрогени, синтезирани при двата пола. Развитието на мъжкия фенотип зависи от Y-хромозомния ген, HY-антигенът (описан при мишката през 1955 г., HA гена). Той се секретира от първични мъжки клетъчни линии. Y 5NA 0 е отговорен за производството на тестостерон (мъжки полов хормон). Веднага след като тези клетки попаднат в началото на половите жлези, започва диференциацията на тестисите. HY рецепторите се намират на повърхността на двата вида гонадни клетки (отклонение от развитието на пола при крави). Смята се, че мъжкият фенотип се определя от цялата мъжка хромозома. Но през 1990 г. генът е открит (Sex Region Y), локализиран в кариотипа на Y хромозомата. В негово отсъствие, XY генотипът дава женски фенотип.
РАЗВИТИЕ НА GONADS
G ┌┬┬┬┐ │ бисексуални зародиши
├┼┼┼┤ │ talnaya
┌────│ └┴┴┴┘ │────┐
│ └───────┘ │
ако генотипът е XX │ │, ако генотипът е XY
(7-8 седмица) ┴──┴──┐ ┌───┴───┐ (6-та седмица)
│ ┌┬┐ │ │┌┬┬┬┬┬┐│
┌─────────┼─├┼┤ │ │├┼┼┼┼┼┼┼────────┐
кортикален ary layer │ │└┴┴┴┴┴┘│ медуларен слой
слой ──────┘ └───────┘
(генът определя женския пол) (генът определя мъжки пол)
При хората специфичните женски хормони, които регулират функциите на
тези признаци стават забележими, те са продукт на интерстициал
нас в края на 7-8 седмици. На 20-та тъкан на яйчника или семенната клетка
първични зародишни клетки на клетката (клетки на Leydig). Те формират
хормонален секс (тестостерон,
гонади и диференцирани до естрадиол). 10-12 седмици за
на oogonyev до края на 2-ри месец. вътрешни сексуални организми
До края на третия месец в дълбочина, на 12-та седмица, със сигурност
гонадните ембриони се откриват на нивото на андрогените в ембриона
ооцити (profase MI). Започва диференциална маскулинизация (
кисти на яйчниците на възраст от 7 месеца (средно изкачване) и завършва
също така ще се стопи до 9-ия месец в яйчника до 20-та седмица.
има 200-400 хиляди ооцити II. В пубертета нивото на est
увеличава и влияе
структура на скелета, както и
съответно андрогени (с
женски и мъжки тип).
Гонадите определят развитието на първични сексуални характеристики и вторично. Сексът жлезите отделят хормони (естрадиоли, андрогени), които заедно с хормоните на ендокринните жлези контролират пътищата на секс диференциация. Нивото на хормоните на свой ред се контролира от гени.
По този начин процесът на сексуална диференциация включва:
1) генетичен контрол;
2) регулаторни функции на хормоните.
Съществува теория за действието на хормоните като регулаторни фактори за гените. Те действат само върху специфични клетки-мишени. В клетката се произвежда специален протеин - рецептор, който се свързва с хормона и същевременно се променя, след което придобива свойствата да индуцира работата на един или няколко гена в хромозомите. Когато глюкокортикоидите действат върху клетките на матката, синтезът на РНК и протеините се променя в тях (виж диаграмата).
Образуването на протеинови рецептори и хормони се контролира от гени. В случай на нарушение на контрола са възможни аномалии, като пример за това е синдромът на Морис. Феминизация на тестисите (синдром на Morris) (Morris, 1953): рецепторите на тестостерон липсват при индивиди с това заболяване. Андрогените се секретират в нормални количества. В синдрома на Морис ембриогенезата е основа за тестисите, които започват да произвеждат мъжки полови хормони. Такива ембриони обаче не образуват рецепторен протеин (рецесивна генна мутация), който осигурява чувствителността на клетките на развиващите се органи към мъжките полови хормони. По този начин развитието на мъжкия тип престава и се появява женският фенотип. В изключителни случаи е възможно такива дефекти да се коригират чрез въвеждане на подходящи хормони.
Така първоначалната генетична бисексуалност е основа за предефиниране на пола. Мъжкият плод придобива характеристиките на женския пол. Мъжки кариотип, мъжки гонади, женски фенотип. Пропорциите на тялото са женски, има млечни жлези, скъсено влагалище, тестиси - в големите срамни устни, ингвинален канал, коремна кухина.
РЕЗОЛЮЦИЯ НА ЕТАЖА
В резултат на излагане на хормони или в резултат на патология на рецепторите на клетката мишена, може да настъпи предефиниране на пола (синдром на Maurice, отстраняване на половите жлези в петлите).
В природата много фактори отслабват действието на гените, които контролират развитието на пола. Например, при хората, в половите жлези на един индивид, тестисите и яйчниците (развиващите се тестиси и яйчници) развиват еднакво хермафродитизъм - феномен на интерсексуалността.
Въз основа на клинични данни има 3 вида интерсекс:
1) истински хермафродитизъм: наличие на зародишни клетки от двата пола;
2) мъжки псевдогермафродитизъм: има само тестиси (тестиси, тестиси), женски фенотип;
3) женски псевдогермафродитизъм: има само яйчници, мъжкия фенотип.
Тази класификация не съвпада с цитогенетичните основи, тъй като 46 XY при човек има опции.
Съотношение на половете
Първичното съотношение на половете (по време на оплождането трябва да бъде близо до 1: 1, тъй като срещата на половите хромозоми е също толкова вероятно). По време на изследването при хора е установено, че на 100 женски зиготи се образуват 140-160 мъжки. Сперматозомите, съдържащи Y хромозомата, са по-леки, по-подвижни и имат голям отрицателен заряд (яйцето има положителен заряд). Ето защо, Y-съдържащите сперматозоиди оплождат яйцеклетката по-често.
Средно - на 100 момичета се раждат 103-105 момчета. Жизнеспособност на женските гамети, чужди протеини на мъжки ембриони. До 20-годишна възраст има 100 момчета на 100 момичета.
Висше - до 50-годишна възраст има 85 мъже на 100 жени, а към 85-годишна възраст има 50 мъже на 100 жени. Женският организъм е по-адаптиран, което може да бъде обяснено, наред с други причини, и от мозаицизма на женския организъм върху половите хромозоми.
Хипотезата на М. Лойон за женския мозаицизъм по отношение на половите хромозоми.
През 1949 г. M. Borr и C. Bertrand установяват, че в ядрата на нервните клетки на жените е открит сноп от интензивно оцветени хроматини. В ядрата на мъжките клетки не се открива. Този сноп се нарича секс хроматин (тялото на Borra) и представлява една инактивирана X хромозома.
В началото на развитието на женския ембрион и двете Х-хромозоми функционират, т.е. два пъти повече от мъжете имат гени. Това може да обясни по-голямата жизнеспособност на женските зиготи.
През 1962 г. М. Лион прави хипотеза за инактивирането на една Х хромозома в женското тяло на бозайници. В женския ембрион и двете хромозоми функционират до ден 16 от ембрионалното развитие. На 16-ия ден, една хромозома се инактивира с образуването на секс хроматин. Този процес е случаен, така че около 1/2 от активните клетки поддържат Х хромозомата на майката X 5М 0, а бащата е инактивиран. В други, бащата е активен (Х 5 0), а майката е инактивирана. Не се извършва повторно активиране. Хромозомите на майката и бащата съдържат алелни, но не съвсем идентични гени, т.е. доминиращ алел е разположен в една хромозома, а другият е рецесивен. Притежаването на допълнителни гени разширява адаптивните способности на организма.
Нива на сексуалната диаграма на сексуалната дифференциация
разграничаване
┌─┐ ┌┴┐
гейм X └ └─┘ Y
яйцеклетка. \\ t
┌───────────────────┐
генетична хромозомия XX и XY │
└───────────────────┘
┌─────────┴─────────┐
гонадална │ неопределена диференцирана
Gon── gonни гонади на ембриона of
│ └───────────────────┘
Ant Y H Y антиген
В периода ┌───────┴───────┐ ┐─────────────┐
генитални gen │ │ │1 генитални │
зреене. │ │ ┌─│ признаци
│ │ │ │ └────────────┘
─────┴─────┐ ┌─────┴─────┐┐ ут in utero
Ond adgondad un adgunad ─────┤ 7-32 седмици
│ │ яйчници │ тестиси ones хормони andro
│ └───────────┘ └───────────┘┘ │ │ клетъчни гени
│ │ Leydinga
Y │ пубертет │ │ ┌────────────┐
└───────────────────────────────┘ │ Диференциация
хормонални хипофизни хормони, мъжки и женски нервни нерви
Ones хормони: андрогени (тестостерон), естро-путейни пътища │
└─ гени (прогистерон) │ пол │
┌──────────────┴───────────────┐ └────────────┘
фенотипна │ Вторични признаци за сексуални характеристики
│└──────────────────────────────┘ │
└──────────────────────────┬────────────────────────┘
┌──────────────────────────┴────────────────────────┐
психологическа and сексуална и поведенческа принадлежност
└───────────────────────────────────────────────────┘
Женското тяло е по-устойчиво на студ, йонизиращо лъчение, емоционални претоварвания (жените плачат по-често, активни амини се открояват със сълзи, в резултат на това кръвното налягане намалява).
Ако хипотезата работи без ограничения, няма да има фенотипни разлики между здрави жени с две X хромозоми и пациенти с X 4 0 или при мъже XY / XXYY. Очевидно, Х-хромозомата не е напълно инактивирана.
Регулиране на съотношението между половете.
Ако поставите сперматозоида в постоянно електрическо поле, ще има разделяне на Х- и Y-хромозомите. Използва се в добитъка. Възможно е да се получат 80% от потомците на желания пол.
Осъзнаването на принадлежността към мъжа или жената е неразделна част от менталното възприятие на пола, психичните параметри. Транссексуализъм - психологически хермафродитизъм., Феноменът на нарушаване на сексуалната идентичност на индивида. Случаи на обсесивно осъзнаване на хората, принадлежащи към противоположния пол, са известни още от древността. Така, Херодот, в своята тайна „Скитска болест” описва човек, който не само носеше женски дрехи, но и възприемаше характерните черти на противоположния пол. Някои императори имаха склонност да се прикриват: например, Калигула. J. D ”Ark.
Несъответствието между анатомичните и умствените параметри е конфликт между вътрешната среда и социалната. В мъжкото тяло живее женската душа и обратно. Направете хирургична корекция на пода. За да превърнете човек в жена, се нуждаете от 1 операция: вагината се формира от кожата на пениса и скротума. От една жена да направи мъж по-труден: 3-4 операции (отстраняване на млечните жлези, формиране на член). Тези хора са обречени на вечни мъки: пият хормони, нямат деца. Институт по човешка репродукция и семейно планиране в Москва.
Сексуалната диференциация включва образуването на първични (гонади) и вторични полови характеристики.
Полът при повечето животни се определя генетично по време на оплождането.
Х-хромозомата носи гена на феминизацията на тестисите (Tfm), нормална
чийто алел определя синтеза на протеиновия рецептор за андрогени,
които се синтезират в женски и мъжки организми.
──────┐ - всяка хромозома е нефункционален Tfm - смисъл
┌┬┬┐ K┌┬┬┐ измамата развива кората.
--────┤ └┴┴┘ ├──────┐┐ XY - съдържа гена I 5HA-ген 0, който е отговорен за
Of └──────┘ │ синтез на антиген, който определя
│ M │ мъж = Отговорен за вземането
│ тестостерон.
┌──┴──┐ ────┴───┐ Диференциацията на половите жлези се наблюдава при 7-10
Of │ │ седмица от живота. На 10-та седмица сексът може да бъде оп
Determine │ │ │ за определяне на набора от хромозоми.
└─────┘ └───────┘
Посоката на развитие се определя от наличието на Y хромозома. Обикновено, ХХ хромозомите съдържат репресорен ген, който предотвратява развитието на мъжки тип. Това развитие зависи от HY (ON) антигена, ген I 5HA е отговорен за производството на тестостерон. Намира се в няколко вида (описани в мишка през 1955 г., Eychwald, Silmser). Той се секретира от първични мъжки клетъчни линии.
Обикновено, при индивиди с комбинация от полови хромозоми, женските гени са доминирани от гените, които определят женския пол, и от мъжки тип доминира мъжкият пол.
Гениталиите се формират от каналите на Мюлер и Волф, които произхождат от основния бъбрек. При жените, каналите на Müller се развиват в фалопиевите тръби и матката, а wolfa тръбите атрофират. При мъжете вълковите канали се развиват в семенните канали и семенните мехурчета. Стероидните хормони тестостерон и дихидротестостерон се синтезират в ембрионални тестиси под влиянието на хормона на майката (хорионгонадотропин). Тези хормони действат върху биопотентните пъпки.
външни и вътрешни генитални органи: вълкови канали, мулерови канали и урогенитални синуси. Нормален мъжки организъм се развива, ако всички тези елементи функционират. В отсъствието им се формират женски половите характеристики. Непълно развитие на мъжкия фенотип с мъжки
генотип (мъжки псевдогермафродитизъм).
Съвременна репродуктивна стратегия:
- изкуствено осеменяване;
- ин витро оплождане;
- изкуствен растеж на ембриона и неговото прехвърляне към матката;
- заместваща майка.
Повечето животни са двудомни организми. Полът може да се разглежда като набор от характеристики и структури, които осигуряват начин за възпроизвеждане на потомство и предаване на наследствена информация. Полът се определя най-често по време на оплождането, т.е. кариотипът на зигота играе основна роля при определяне на пола. Кариотипът на всеки организъм съдържа хромозоми, които са еднакви и при двата пола - автозоми, и хромозоми, при които женският и мъжкият пол се различават един от друг - половите хромозоми. При хората, "женските" полови хромозоми са две Х-хромозоми. По време на образуването на гамети всяко яйце получава един от Х хромозомите. Полът, в който се формират гамети от същия тип, които носят Х-хромозомата, се нарича хомогаметик. При хората женският пол е хомогаметичен. "Човешки" полови хромозоми при хора са Х хромозомата и Y хромозомата. По време на образуването на гамети половината от сперматозоидите получават Х-хромозомата, а другата половина Y-хромозома. Етаж, който има различни видове гамети се нарича хетерогамет. При хората мъжкият пол е хетерогамет. Ако се образува зигота, която носи две Х-хромозоми, тогава от нея ще се образува женско тяло, ако Х-хромозомата и Y-хромозомата са мъжки.
При животните могат да се различат следните четири вида хромозомално определяне на пола.
1. Женският пол е хомогаметичен (XX), мъжкият пол е хетерогамет (XY) (бозайници, по-специално, човешка, плодова муха).
Генетична схема за определяне на хромозомния пол при хора:
Генетична схема за определяне на хромозомния пол в дрозофила:
2. Женският пол е хомогаметичен (ХХ), мъжкият пол е хетерогаметен (Х0) (ортопери).
Генетична схема за определяне на хромозомния пол в пустинята:
3. Женският пол е хетерогамет (XY), мъжкият пол е хомогаметичен (XX) (птици, влечуги).
Генетична схема за определяне на хромозомния пол в гълъба:
Yandex.DirectВсички реклами
4. Женският пол е хетерогаметен (Х0), мъжкият пол е хомогаметичен (ХХ) (някои видове насекоми).
Определяне на пола при хора се случва с механизма XY. В същото време хетерогаметният пол е мъжки, хомогаметичен - женски. Определянето на пола се разделя на три етапа: хромозомна, гонадна и фенотипна.
Две основни правила за определяне на пола при бозайници
Класическите ембриогенетични изследвания са установили две правила за определяне на пола при бозайници. Първият от тях е формулиран през 1960-те от Алфред Йост въз основа на експерименти за премахване на зародиша на бъдещите гонади (гонаден валяк) в ранните зайци от зайци: премахването на ролките преди образуването на гонадите доведе до развитието на всички ембриони като женски, Предполага се, че гонадите на мъжкия ефекторен хормон тестостерон, отговорен за маскулинизацията на фетусите, се секретират и се предвижда наличието на втори ефектор на анти-Мюлер хормон (MIS), който директно контролира такива анатомични трансформации. Резултатите от наблюденията са формулирани по правило: специализацията на развиващите се гонади в тестисите или яйчниците определя последващата сексуална диференциация на ембриона.
До 1959 г. се приема, че броят на Х-хромозомите е най-важният фактор, контролиращ пола при бозайници. Въпреки това, откритието на организми с един Х-хромозома, развиващо се като женски, и индивиди с една Y хромозома и множество Х хромозоми, които се развиват като мъже, ги принуждава да се откажат от тези понятия. Формулирано е второто правило за определяне на пола при бозайници: Y-хромозомата носи генетичната информация, необходима за определяне на пола при мъжете.
Комбинацията от горните две правила понякога се нарича принцип на растеж: Хромозомният пол, свързан с наличието или отсъствието на Y хромозомата, определя диференциацията на ембрионалните гонади, която от своя страна контролира фенотипния пол на организма. Такъв механизъм за определяне на пола се нарича генетичен. GSD) и се противопоставят на такива, въз основа на контролиращата роля на факторите на околната среда (англ. ESD) или съотношението на половите хромозоми и автозомите (eng.). CSD).
Определяне на хормоналния пол
Дефиницията на пола може да бъде представена като палка, която хромозомният механизъм предава на недиференцирани гонади, които се развиват в мъжки или женски полови органи. При изследване на ролята на половите хромозоми в развитието на половите жлези е показано, че наличието или отсъствието на Y хромозомата е решаващо за хората. При отсъствието на Y хромозомата възниква диференциране на гонадите в яйчниците и се развива жена. В присъствието на Y хромозомата, мъжката система се развива. Очевидно, Y хромозомата произвежда вещество, което стимулира тестикуларната диференциация. "Изглежда, че основният план на природата е да се направи жена, и че добавянето на Y-хромозома произвежда човек-вариация." Следващият етап от релето продължава хормоните, които определят процеса на сексуална диференциация на плода и неговото анатомично развитие. При раждането се завършва първата част от програмата. След раждането палката се придвижва към екологични фактори, които завършват формирането на пола - обикновено, но не винаги, според генетичния пол. Определянето на пола е сложен многостепенен процес, който при хората зависи, освен от биологичните, и от психосоциалните фактори. Това може да доведе до транссексуално, хетеросексуално, бисексуално или хомосексуално поведение и начин на живот.
Физиологична основа за определяне на пола на половите жлези
Физиологичната основа на механизма за определяне на пола е бисексуалността на феталните гонади на бозайници. В такъв progonadah едновременно представя Мюлеров канал и канал Wolff - начало на гениталния тракт от мъже и жени, респективно. Първичното определяне на пола започва с появата на специализирани клетъчни линии в прогоните - клетката Sertoli. В миналото синтезира прогнозира Йост antimyulllerovsky хормон, отговорен за пряка или непряка инхибиране на Мюлеров канал - на зачатък на бъдещето на маточните тръби и матката на.
Генетичен механизъм за определяне на пола
Човешка Y хромозома, показваща местоположението на SRY ген
През 1987, David Page и колегите му проучват мъж XX, наследена специфичен фрагмент от дължината на Y-хромозома на 280,000 базови двойки и XY жената с делеция, тази широка площ, в резултат на обмена между хромозоми секции. Този фрагмент присъства в Y-хромозома на всички тези животни Eutheria и на разстояние 100 килобази от граничната област ген psevdoautosomalnoy ZFY дължина от 140 килобази.
ZFY хомологът - ZFX генът се намира в X хромозомата и ZFX не е инактивиран. И двата фактора ZFX и ZFY кодират транскрипционни фактори, съдържащи мотиви на цинкови пръсти с ДНК-свързваща активност. По-нататъшният подробен анализ на специфични последователности на Y-хромозоми при индивиди с инверсия на секса ограничи търсенето до площ от 35 kbp по размер и доведе до откриването на ген, считан за истински еквивалент на класическия английски. Определящ тест тест, Този ген се нарича SRY (eng.). Гени за определяне на пола в регион Y).
КРЗ намираща се в областта на определяне на пола и съдържа консервативен домен (HMG-box), кодиращ протеин от 80 аминокиселинни остатъка. КРЗ генна активност, наблюдавана преди началото на периода на диференциация progonady в яйчен - 10-12-ия ден на ембрионалното развитие при мишки и най-малко на този етап не зависи от присъствието на зародишни клетки. Специфичните точкови мутации или делеции в HMG кутията на този ген в XY жени водят до инверсия на пола. Прехвърляне на ДНК фрагмент от 14 килобазисни двойки, съдържащ ген с фланкиращи региони, оплодената яйцеклетка homogametic лица чрез микроинжектиране доведе до появата на мъжки кариотип с XX.
SRY генни функции
Домейнът, кодиран от HMG кутията на SRY гена, се свързва специфично с ДНК и я кара да се огъва. Огъване на ДНК индуциран протеин КРЗ или негови хомолози, съдържащи домен на HMG, може да бъде механично предадени на значително разстояние, и играят важна роля в регулацията на транскрипцията, репликация и рекомбинация. ОБЛАСТ ДНК, в която локализиран КРЗ, съдържа два гена, кодиращи ключови ензими, включени в диференциацията на първични полови жлези на мъжки тип: ген ароматаза Р450 контролиране на превръщането на тестостерон в естрадиол и фактор инхибиране развитието Miller канали, което води до обръщане на тяхното развитие и насърчава диференциацията на тестисите ,
КРЗ генен продукт, също се включва в процеса на сексуалната диференциация тясно с друг ген, наречен геном Z, чиято функция е нормално мъжки специфично инхибиране на гени. В случай на нормално мъжки генотип 46XY КРЗ ген кодира протеин, натискане Z ген, и специфични мъжки гени са активирани. В случай на нормална женска генотип 46HH в които няма КРЗ, Z ген се активира и инхибира специфичен мъжки ген, който създава условия за развитие на женски тип.
Хромозомна теория за определяне на пола
Помислете за определяне на хромозомния пол. Известно е, че в двудомни организми (животни и двудомни растения) съотношението между половете обикновено е 1: 1, т.е. мъжете и жените са еднакво разпространени. Това съотношение съвпада с разделянето в анализиращия кръст, когато една от кръстосаните форми е хетерозиготна (Аа), а другият е хомозиготен за рецесивните алели (Аа). В потомството в този случай има разделяне по отношение на 1 Аа: 1аа. Ако даден пол се наследява по същия принцип, би било логично да се предположи, че един пол трябва да бъде хомозиготен, а другият хетерозиготен. Тогава разделянето по пол трябва да бъде равно на 1.1 във всяко поколение, което се наблюдава.
Еволюционна теория на пола В. Геодакян е предложен през 1965 г. Теорията обяснява от единна гледна точка много явления, свързани с пола: сексуален диморфизъм в здравето и болестите, съотношение между половете, диференциална смъртност и сексуален отговор, ролята на половите хромозоми, половите хормони, асиметрията на мозъка и ръката, реципрочни ефекти и психологически и социални различия между половете.
Теорията се основава на принципа на спрегнатите подсистеми, които се развиват асинхронно. Мъжът е оперативен подсистема на населението, жени - консервативен подсистема. Новата информация от околната среда първо достига мъжкия пол и едва след като много поколения се прехвърлят на жената, еволюцията на мъжкия пол предхожда еволюцията на женската. Тази смяна на времето (две фаза развитие на черта) създава две форми на черта (мъжки и женски) - сексуален диморфизъм в популацията. Необходима е еволюционна “дистанция” между подсистемите за търсене и проверка на иновациите.
 Най-страшното заболяване в света
Най-страшното заболяване в света Пълно и непълно господство
Пълно и непълно господство Как можете да направите приятелката си щастлива
Как можете да направите приятелката си щастлива