История генетики человека этапы развития науки кратко. Основные даты в истории генетики
Введение
Биологические системы обладают способностью сохранять и передавать информацию в виде структур и функций, возникших в прошлом в результате длительной эволюции.
Открыты подвижные генетические элементы, которые оказались замешаны в таких общебиологических явлениях, как азотфиксация, злокачественный рост клеток, работа иммунной системы и приспособление бактерий к антибиотикам, нестабильные мутации, материнская наследственность.
Генетика - это биологическая наука о наследственности и изменчивости организмов и методах управления ими.
Центральным понятием генетики является "ген". Это элементарная единица наследственности, характеризующаяся рядом признаков.
В основу генетики легли закономерности наследственности, обнаруженные австрийским биологом Г. Менделем при проведении им серии опытов по скрещиванию различных сортов гороха.
Основные направления исследований учёных - генетиков:
1. Изучение молекул нуклеиновых кислот, являющихся хранителями генетической информации каждого вида живого, единицами наследственности.
2. Исследование механизмов и закономерностей передачи генетической информации.
3. Изучение механизмов реализации генетической информации в конкретные признаки и свойства живого.
4. Выяснение причин и механизмов изменения генетической информации на разных этапах развития организма.
> История развития генетики как науки
После повсеместного распространения учения Ч. Дарвина одним из первых критиков, указавших на слабое место в теории, был шотландский исследователь Ф. Дженкинс. В 1867 г. он заметил, что в дарвиновской теории нет ясности в вопросе о том, как осуществляется накопление в потомстве тех или иных изменений. Ведь сначала изменения признака происходят только у некоторых особей. После скрещивания с нормальными особями должно наблюдаться не накопление, а разбавление данного признака в потомстве. То есть в первом поколении остаётся? изменения, во втором - ? изменения и т. д. вплоть до полного исчезновения этого признака. Ч. Дарвин так и не нашёл ответа на этот вопрос.
Между тем решение этого вопроса существовало. Его получил преподаватель монастырской школы в Брно (Чехия) Г. Мендель. В 1865 году были опубликованы результаты его работ по гибридизации сортов гороха, где были открыты важнейшие законы наследственности. Автор показал, что признаки организмов определяются дискретными наследственными факторами.
Он ещё до выхода в свет книги Ч. Дарвина хотел проследить судьбу изменений генотипов в разных поколениях гибридов. Объектом исследования стал горох. Мендель взял два сорта гороха - с жёлтыми и с зелёными семенами. Скрестив эти два сорта, он обнаружил в первом поколении гибридов горох только с жёлтыми семенами. Путём самоопыления полученных гибридов он получил второе поколение. В нём появились особи с зелёными семенами, но их было заметно меньше, чем с жёлтыми. Подсчитав число тех и других, Мендель пришёл к выводу, что число особей с желтыми семенами относится к числу особей с зелёными как приблизительно 3:1.
Параллельно он проводил серию других опытов с растениями, прослеживая какой-либо признак в нескольких поколениях. В каждом опыте в первом поколении проявлялся только один из родительских признаков. Мендель назвал его доминантным. Временно исчезающий признак он назвал рецессивным. Во всех опытах отношение числа особей с доминантным признаком к числу особей с рецессивным признаком среди гибридов второго поколения было в среднем равно 3:1.
Итак, можно было утверждать, что при скрещивании растений с противоположными признаками происходит не разбавление признаков, а подавление одного признака другим, в связи с этим необходимо различать доминантные и рецессивные признаки.
Мендель пошёл в своих экспериментах дальше. Он произвёл самоопыление гибридов второго поколения и получил гибриды третьего, а затем и четвёртого поколения. Он обнаружил, что гибриды второго поколения с рецессивным признаком при дальнейшем размножении не расщепляются ни в третьем, ни в четвёртом поколениях. Так же ведёт себя примерно треть гибридов второго поколения с доминантным признаком. Две трети гибридов с доминантным признаком расщепляются при переходе к гибридам третьего поколения, причём опять-таки в отношении 3:1. Получившиеся при этом расщеплении гибриды третьего поколения с рецессивным признаком и треть гибридов с доминантным признаком при переходе к четвёртому поколению не расщепляются, а остальные гибриды третьего поколения расщепляются, причём снова в отношении 3:1.
Этот факт демонстрирует важное обстоятельство: особи с одинаковыми внешними признаками могут обладать разными наследственными свойствами, то есть по фенотипу нельзя судить с достаточной полнотой о генотипе. Если особь не обнаруживает в потомстве расщепления, то её называют гомозиготной, если обнаруживает - гетерозиготной.
В итоге Г. Менделем был сформулирован закон единообразия гибридов первого поколения: первое поколение гибридов в силу проявления у них лишь доминантных признаков всегда единообразно. Этот закон носит также название первого закона Менделя или закона доминирования. Однако результаты его исследований оставались практически неизвестными почти 35 лет - с 1865 по 1900.
В 1900 году законы Менделя были переоткрыты независимо сразу тремя учеными - Г. де Фризом в Голландии, К. Корренсом в Германии и Э. Чермаком в Австрии. В 1909 датский ученый В. Иогансен ввёл понятие «ген» (от греч. слова «происхождение»).
Хромосомная теория наследственности, разработанная в 1910-1915 годах в трудах А. Вейсмана, Т. Моргана, А. Стертеванта, Г. Дж. Меллера и других утверждает, что передача признаков и свойств организма от поколения к поколению (наследственность) осуществляется в основном через хромосомы, в которых расположены гены.
В 1944 году американскими биохимиками (О. Эвери и др.) было установлено, что носителем свойства наследственности является ДНК.
С этого времени началось быстрое развитие науки, исследующей основные проявления жизни на молекулярном уровне. Тогда же впервые появился новый термин для обозначения этой науки - молекулярная биология. Молекулярная биология исследует, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и другие явления обусловлены структурой и свойствами биологически важных молекул (главным образом белков и нуклеиновых кислот).
В 1953 году была расшифрована структура ДНК (Ф. Крик, Д. Уотсон). Расшифровка структуры ДНК показала, что молекула ДНК состоит из двух комплементарных полинуклеотидных цепей, каждая из которых выступает в качестве матрицы для синтеза новых аналогичных цепей. Свойство удвоения ДНК обеспечивает явление наследственности.
Расшифровка структуры ДНК была революцией в молекулярной биологии, которая открыла период важнейших открытий, общее направление которых - выработка представлений о сущности жизни, о природе наследственности, изменчивости, обмена веществ. В соответствии с молекулярной биологией, белки - это очень сложные макромолекулы, структурными элементами которых являются аминокислоты. Структура белка задается последовательностью образующих его аминокислот. При этом из 100 известных в органической химии аминокислот в образовании белков всех организмов используется только двадцать. До сих пор не ясно, почему именно эти 20 аминокислот синтезируют белки органического мира. Вообще, в любом существе, живущем на Земле, присутствуют 20 аминокислот, 5 оснований, 2 углевода и 1 фосфат.
К концу XIX века в результате повышения оптических качеств микроскопов и совершенствования цитологических методов стало возможно наблюдать поведение хромосом в гаметах и зиготах.
Материальные основы наследственности стали проясняться около 50 лет назад, когда Ф. Крик и Дж. Уотсон расшифровали строение ДНК. Задолго до этого биологи, изучая передачу наследственных признаков при скрещивании, поняли, что каждый признак определяется отдельной частицей, которую назвали геном. Оказалось, что гены лежат в ядре клетки, в хромосомах. После открытия роли ДНК и механизма синтеза белков стало ясно, что ген - это участок цепочки ДНК, на котором записано строение молекулы определённого белка. В некоторых генах всего 800 пар нуклеотид, в других - около миллиона. У человека всего около 90 тыс. генов.
Каждая прядь молекулы ДНК представляет собой цепочку из четырёх типов звеньев - нуклеотид, повторяющихся в разном порядке. Нуклеотиды обычно считают парами, так как в молекуле ДНК две цепочки и их нуклеотиды соединены поперечными связями попарно. Четыре сорта нуклеотид, четыре «буквы» позволяют записать генетический текст, который прочитывается механизмами синтеза белка в живой клетке. Группа из трёх стоящих подряд нуклеотид, действуя через довольно сложный механизм передачи, заставляет рибосому - внутриклеточную частичку, занимающуюся синтезом белков - подхватывать из цитоплазмы определённую аминокислоту. Затем следующие три нуклеотида через посредников диктуют рибосоме, какую аминокислоту ставить в цепочку белка на следующее место и так постепенно получается молекула белка. Информации, записанной в ДНК тройками пар нуклеотид, достаточно для построения нового организма со всеми его особенностями.
Генетическая информация хранится в виде последовательности нуклеотид. Она передаётся в клетке от ДНК к РНК. В процессе этой реакции воспроизводится часть последовательности ДНК, ген и синтезируется матричная РНК. Последовательность матричной РНК, состоящей только из одной цепи, является комплементарной последовательности нуклеотид кодирующей её цепи ДНК.
Зародилась новая отрасль генетики - геномика, изучающая целые геномы. До недавнего времени на основе достижений молекулярной биологии и генной инженерии удалось прочитать генетические тексты вирусов грибков, дрожжевых бактерий и, наконец, в 1998 году после 8 лет напряженной работы удалось прочитать геном многоклеточного животного - нематоды (маленького червячка, обитающего в почве). Расшифрован геном человека. Геном нематоды состоит из примерно 100 миллионов пар нуклеотид. Геном человека состоит из 3 миллиардов пар. Создана международная программа «Геном человека». Лаборатории в разных странах сообщают данные о расшифровке нуклеотид (секвенировании) в Международный банк данных, доступных каждому исследователю.
Ее результаты существенны для понимания происхождения человека и других видов, эволюции молекул и клеток, взаимодействия информации с потоками веществ и энергии в живых системах. Сегодня ученые полностью расшифровали структуру и расположение всех генов, присутствующих в человеческом организме. Но потребуется значительное время и средства, чтобы понять законы функционирования генов - партитуру, которая превращает солистов (гены) в слаженный оркестр.
Биогенетический закон сформулировал Э. Геккель в 1866 г. на основе идей Ч. Дарвина и исследований Ф. Мюллера. Поэтому он носит название биогенетического закона Э. Геккеля - Ф. Мюллера. Биогенетический закон в 1910 г. существенно уточнил А. Н. Северцев (1866 - 1936), создавший теорию филэмбриогенеза.
Согласно этому закону, зародыши в процессе развития повторяют в несколько сокращённом виде эволюционный путь, пройденный их предками, то есть существует сходство между эмбриональным развитием и эволюционным процессом. В настоящее время установлено, что зародыши высших форм животных сходны с зародышами низших форм. Ранние стадии развития зародыша удивительно сходны у всех позвоночных, и нелегко отличить зародыш человека от зародыша свиньи, цыплёнка, лягушки или рыбы. Повторение (рекапитуляция) в онтогенезе филогенетических черт может быть неполным, с определенными искажениями, связанными с дальнейшими эволюционными преобразованиями, в частности могут повторяться особенности соответствующих фаз развития предковых форм.
Важнейшее достижение биогенеза заключается в формировании генетических программирующих устройств, позволяющих закреплять достигнутое. Соревнование различных программ в борьбе за существование ведёт к двум важным следствиям:
Во-первых, естественный отбор совершенствует программы индивидуального развития особей.
Во-вторых, возникает программирование направления эволюции видов. При этом программирующим устройством становится сама биосфера. Ведь она определяет особенности, скорость и направление эволюционных преобразований видов, входящих в её состав.
История генетики как науки началась не так уж давно, примерно со второй половины XIX века, с работ чешского монаха Грегора Менделя. До этого люди не знали, что лежит в основе наследственности и изменчивости организмов. Никаких систематических исследований в этой области не проводилось. С древних времен существовали две гипотезы о том, почему потомки похожи на своих предков. Это гипотезы прямого и непрямого наследования.
По гипотезе прямого наследования предполагалось, что каждая часть тела родителя, каждая его клетка передает особенности своего строения потомку. Гипотеза непрямого наследования исходила из того, что не все части организма участвуют в образовании половых продуктов, а они образуются обособлено, поэтому прижизненные изменения родителя не передаются потомкам.
Чарльз Дарвин придерживался первой гипотезы, что было ошибкой. Из-за этого он не смог опровергнуть одного из критиков своей эволюционной теории, который писал следующее. Если у единичного животного появляется какой-либо признак, то при передаче следующему поколения от него останется ½, еще следующему ¼ и т. д. В конечном итоге признак исчезнет. Дарвин не смог возразить, так как не знал, что признаки организма дискретны, они не смешиваются и не растворяются.
Г. Мендель провел длительные, систематические, статистические исследования наследования признаков. В качестве объекта исследования был выбран горох. Это был очень удачный выбор. Такие признаки как цвет семян и цветков, морщинистость семян и ряд других находятся в разных хромосомах (т. е. наследуются не сцеплено), и у них чаще всего нет промежуточного проявления признака (например, семя может быть либо желтым, либо зеленым, но не смешанного цвета). Мендель конечно же тогда ничего не знал о сцепленном наследовании, влиянии генов и аллелей друг на друга и др. Если бы в исследуемых признаках гороха все это наблюдалось, то его эксперимент бы мог не удасться.
Грегор Мендель и открытые им законы наследственности - 1860-е годы
Мендель доказал дискретность наследственных признаков. Они не разбавляются, а существует лишь подавление одного другим. Менделем был разработан гибридологический метод исследования. Но самое главное Мендель впервые в истории генетики сформулировал : единообразия гибридов первого поколения, расщепления во втором поколении, независимого наследования .
Однако в те времена история генетики еще не началась. Мендель был монахом-самоучкой, и его исследованиям большого значения не придали. Лишь в начале XX века, когда многие ученые экспериментально на разных растениях и животных подтвердили справедливость законов Менделя, его работы получили заслуженную оценку.
Начало XX века было бурным этапом в развитии генетики. В это время появляется сам термин «генетика». Дается определение «гена», «генотипа» и «фенотипа». Обнаруживается явление (совместного наследования) генов, У. Бэтсоном открывается и др. В 1910 году Т. Морган совместно с другими учеными разрабатывает хромосомную теорию, которая во многом обобщает и объясняет все ранее сделанные открытия в истории генетики.
 Томас Морган изучал хромосомы, открыл кроссинговер - 1910-е годы
Томас Морган изучал хромосомы, открыл кроссинговер - 1910-е годы
В последующие годы генетика и эволюционное учение связываются между собой. Второе находит объяснение с точки зрения законов первой.
Ученые знали, что хромосомы участвуют в передаче наследственной информации, но не знали, какое вещество ответственно за это. В 40-х годах стало понятно, что ДНК является носителем наследственности. Так ряд ученых переносили ДНК одних бактерий в другие и наблюдали у вторых появление признаков первых.
С развитием методов химии и физики стало возможно исследовать структуру ДНК, что было сделано Криком и Уотсоном в 1953 году. Оказалось, что молекула состоит из двух полинуклеотидных цепей, закрученных в спираль. Каждая из цепей ДНК является матрицей для синтеза комплементарной ей новой цепи, и удвоение ДНК обеспечивает наследственность.
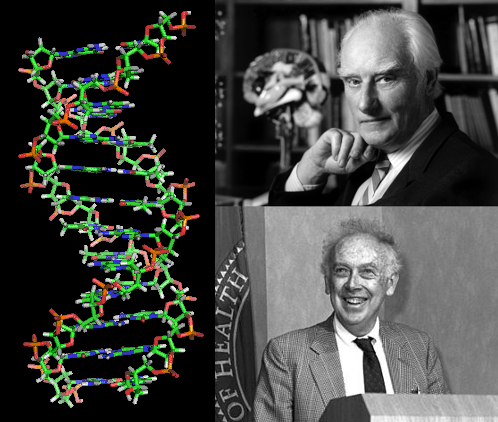 Фрэнсис Крик и Джеймс Уотсон открыли структуру ДНК - 1950-е годы
Фрэнсис Крик и Джеймс Уотсон открыли структуру ДНК - 1950-е годы
Ученые поняли, что последовательность нуклеотидов в гене определяет структуру белковой молекулы. .
В истории генетики 70-е годы XX века ознаменовались появлением генной инженерии. Ученые стали вмешиваться в геномы живых организмов и изменять их. Стали изучаться молекулярные основы различных физиологических процессов.
В последнее десятилетие XX века были секвенированы (расшифрованы) геномы многих простых организмов. В начала XXI века (2003 г.) был завершен проект по расшифровке (определению последовательности нуклеотидов в хромосомах) генома человека.
 Логотип проекта "Геном человека"
Логотип проекта "Геном человека"
На сегодняшний день существуют базы данных геномов многих организмов. Наличие такой базы данных человека имеет большое значение в предупреждении и исследовании многих заболеваний.
Рождение генетики на рубеже двух веков (1900) было подготовлено всем предшествующим развитием биологической науки. XIX в. вошел в историю биологии благодаря двум великим открытиям: клеточной теории, сформулированной М. Шлейденом и Т. Шванном (1838), и эволюционному учению Ч. Дарвина (1859). Оба открытия сыграли определяющую роль в становлении генетики. Клеточная теория, объявившая клетку основной структурной и функциональной единицей всех живых существ, вызвала повышенный интерес к изучению ее строения, что в дальнейшем привело к открытию хромосом и описанию процесса клеточного деления. В свою очередь, теория Ч. Дарвина касалась важнейших свойств живых организмов, которые стали впоследствии предметом изучения генетики — наследственности и изменчивости. Обе теории в конце XIX в. объединила идея о необходимости существования материальных носителей этих свойств, которые должны находиться в клетках.
До начала ХХ в. все гипотезы о механизмах наследственности носили чисто умозрительный характер. Так, согласно теории пангенезиса Ч. Дарвина (1868) от всех клеток организма отделяются мельчайшие частицы — геммулы, которые циркулируют по кровяному руслу и попадают в половые клетки. После слияния половых клеток, в ходе развития нового организма, из каждой геммулы образуется клетка того же типа, от которого она произошла, обладающая всеми свойствами, в том числе и приобретенными родителями в течение жизни. Корни воззрения Дарвина относительно механизма передачи признаков от родителей к потомству через кровь лежат еще в натурфилософии древнегреческих философов, в том числе в учении Гиппократа (V в. до н.э.).
Еще одна умозрительная гипотеза наследственности была выдвинута в 1884 г. К. Негели (нем.). Он предположил, что в передаче наследственных задатков потомству принимает участие особое вещество наследственности — идиоплазма, состоящая из молекул, собранных в клетках в крупные нитевидные структуры — мицеллы. Мицеллы соединяются в пучки и образуют сеть, которая пронизывает все клетки. Идиоплазмой обладают как половые, так и соматические клетки. Остальная часть цитоплазмы в передаче наследственных свойств участия не принимает. Не будучи подкреплена фактами, гипотеза К. Негели, тем не менее, предвосхитила данные о существовании и структурированности материальных носителей наследственности.
Впервые на хромосомы как материальные носители наследственности указал А. Вейсман. В своей теории он исходил из выводов немецкого цитолога Вильгельма Ру (1883) о линейном расположении в хромосомах наследственных факторов (хроматиновых зерен) и продольном расщеплении хромосом во время деления как возможном способе распределения наследственного материала. Теория “зародышевой плазмы” А. Вейсмана получила окончательное оформление в 1892 г. Он считал, что в организмах существует особое вещество наследственности — “зародышевая плазма”. Материальным субстратом зародышевой плазмы являются хроматиновые структуры ядер половых клеток. Зародышевая плазма бессмертна, через половые клетки она передается потомкам, тогда как тело организма — сома — является смертным. Зародышевая плазма состоит из дискретных частиц — биофор, каждая из которых определяет отдельное свойство клеток. Биофоры группируются в детерминанты — частицы, определяющие специализацию клеток. Они, в свою очередь, объединяются в структуры более высокого порядка (иды), из которых формируются хромосомы (по терминологии А. Вейсмана —).
А. Вейсман отрицал возможность наследования приобретенных свойств. Источником наследственных изменений, согласно его учению, служат события, которые происходят в ходе процесса оплодотворения: потеря части информации (редукция) во время созревания половых клеток и смешение детерминантов отца и матери, приводящее к появлению новых свойств. Теория А. Вейсмана оказала огромное влияние на развитие генетики, определив дальнейшее направление генетических исследований.
К началу ХХ в. были созданы реальные предпосылки для развития генетической науки. Решающую роль сыграло переоткрытие в 1900 г. законов Г. Менделя. Чешский исследователь-любитель, монах Брюннского монастыря Грегор Мендель еще в 1865 г. сформулировал основные законы наследственности. Это стало возможным благодаря разработке им первого научного генетического метода, который получил название “гибридологического”. В его основу была положена система скрещиваний, позволяющая вскрывать закономерности наследования признаков. Менделем были сформулированы три закона и правило “чистоты гамет”, которые будут подробно рассмотрены в следующей лекции. Не менее (а, может быть, более) важным было то, что Мендель ввел понятие о наследственных задатках (прообразах генов), которые служат материальной основой развития признаков, и высказал гениальную догадку об их парности как результате слияния “чистых” гамет.
Исследования Менделя и его взгляды на механизм наследования опередили развитие науки на несколько десятилетий. Даже умозрительные гипотезы о природе наследственности, о которых говорилось выше, были сформулированы позже. Еще не были открыты хромосомы и не был описан процесс клеточного деления, который лежит в основе передачи наследственной информации от родителей к потомкам. В связи с этим современники, даже те, кто подобно Ч. Дарвину был знаком с работами Г. Менделя, не сумели по достоинству оценить его открытие. На протяжении 35 лет оно не было востребовано биологической наукой.
Справедливость восторжествовала в 1900 г., когда последовало вторичное переоткрытие законов Менделя одновременно и независимо тремя учеными: Г. де Фризом (голл.), К. Корренсом (нем.) и Э. Чермаком (австр.). Повторив эксперименты Менделя, они подтвердили универсальный характер открытых им закономерностей. Менделя стали считать основателем генетики, и с 1900 г. начался отсчет развития этой науки.
В истории генетики обычно выделяют два периода: первый — период классической, или формальной, генетики (1900-1944) и второй — период молекулярной генетики, который продолжается до настоящего времени. Основная особенность первого периода заключается в том, что природа материальных носителей наследственности оставалась неизвестной. Введенное датским генетиком В. Иогансеном понятие “ген” — аналог менделевского наследственного фактора — было абстрактным. Вот цитата из его работы 1909 г.: “Свойства организма обусловливаются особыми, при известных обстоятельствах отделимыми друг от друга и в силу этого до известной степени самостоятельными единицами или элементами в половых клетках, которые мы называем генами. В настоящее время нельзя составить никакого определенного представления о природе генов, мы можем лишь довольствоваться тем, что подобные элементы действительно существуют. Но являются ли они химическими образованиями? Об этом мы пока не знаем решительно ничего”. Несмотря на отсутствие знаний о физико-химической природе гена, именно в этот период были вскрыты основные законы генетики и разработаны генетические теории, составившие фундамент этой науки.
Переоткрытие законов Менделя в 1900 г. привело к быстрому распространению его учения и многочисленным, чаще всего успешным, попыткам исследователей в разных странах на разных объектах (куры, бабочки, грызуны и др.) подтвердить универсальный характер его законов. В ходе этих экспериментов были вскрыты новые закономерности наследования. В 1906 г. английские ученые У. Бэтсон и Р. Пеннет описали первый случай отклонения от законов Менделя, названный позже сцеплением генов. В этом же году английский генетик Л. Донкастер в опытах с бабочкой обнаружил явление сцепления признака с полом. Одновременно в начале ХХ в. начинается изучение стойких наследственных изменений мутаций (Г. де Фриз, С. Коржинский), а также появляются первые работы по генетике популяций. В 1908 г. Г. Харди и В. Вайнберг сформулировали основной закон генетики популяций о постоянстве частот генов.
Но наиболее важными исследованиями периода классической генетики были работы выдающегося американского генетика Т. Моргана и его учеников. Т. Морган является основателем и руководителем крупнейшей в мире генетической школы, из которой вышла целая плеяда талантливых генетиков. В своих исследованиях Морган впервые использовал плодовую мушку дрозофилу, которая стала излюбленным генетическим объектом и продолжает им оставаться и сейчас. Изучение явления сцепления генов, открытого У. Бетсоном и Р. Пеннетом, позволило Моргану сформулировать основные положения хромосомной теории наследственности, с которыми мы подробно познакомимся ниже. Главный тезис этой базовой генетической теории заключался в том, что гены в линейном порядке располагаются в хромосоме, подобно бусинкам на ниточке. Однако даже в 1937 г. Морган писал о том, что среди генетиков нет согласия в точке зрения на природу гена — являются ли они реальными или абстракцией. Но отмечал, что в любом случае ген ассоциирован со специфической хромосомой и может быть локализован там путем чистого генетического анализа.
Морганом и его коллегами (Т. Пайнтер, К. Бриджес, А. Стертевант и др.) выполнен ряд других выдающихся исследований: разработан принцип генетического картирования, создана хромосомная теория определения пола, изучена структура политенных хромосом.
Важным событием периода классической генетики было развитие работ по искусственному мутагенезу, первые данные о котором были получены в 1925 г. в СССР Г.А. Надсоном и Т.С. Филипповым в опытах по облучению дрожжевых клеток радием. Решающее значение для развертывания работ в этом направлении имели эксперименты американского генетика Г. Меллера по воздействию рентгеновских лучей на дрозофилу и разработка им методов количественного учета мутаций. Работа Г. Меллера вызвала огромное число экспериментальных исследований с использованием рентгеновских лучей на разных объектах. В результате был установлен их универсальный мутагенный эффект. Позже было обнаружено, что мутагенным действием обладают и другие типы излучения, например УФ, а также высокая температура и некоторые химические вещества. Первые химические мутагены были открыты в 30-х гг. в СССР в экспериментах В.В. Сахарова, М.Е. Лобашева и С.М. Гершензона и их сотрудников. Через несколько лет это направление приобрело широкий размах, особенно благодаря исследованиям А.И. Рапопорта в СССР и Ш. Ауэрбаха в Англии.
Исследования в области экспериментального мутагенеза привели к быстрому прогрессу в познании мутационного процесса и к выяснению ряда вопросов, касающихся тонкой структуры гена.
Еще одно важное направление генетических исследований в период классической генетики касалось изучения роли генетических процессов в эволюции. Основополагающие работы в этой области принадлежат С. Райту, Р. Фишеру, Дж. Холдейну и С.С. Четверикову. Своими трудами они подтвердили правильность основных положений дарвинизма и способствовали созданию новой современной синтетической теории эволюции, которая представляет собой результат синтеза теории Дарвина и генетики популяций.
С 1940 г. начался второй период в развитии мировой генетики, который получил название молекулярного, в соответствии с лидирующим положением этого направления генетической науки. Основную роль в бурном подъеме молекулярной генетики сыграл тесный альянс биологов с учеными других областей естествознания (физики, математики, кибернетики, химии), на волне которого был сделан ряд важнейших открытий. В течение этого периода ученые установили химическую природу гена, определили механизмы его действия и контроля и сделали еще много важнейших открытий, которые превратили генетику в одну из основных биологических дисциплин, определяющих прогресс современного естествознания. Открытия молекулярной генетики не опровергли, а лишь вскрыли глубинные механизмы тех генетических закономерностей, которые были вскрыты формальными генетиками.
Работами Дж. Бидла и Э. Тетума (США) было установлено, что мутации у хлебной плесени Neurospora crassa блокируют различные этапы клеточного метаболизма. Авторы высказали предположение, что гены контролируют биосинтез ферментов. Появился тезис: “один ген — один фермент”. В 1944 г. исследование по генетической трансформации у бактерий, выполненное американскими учеными (О. Эйвери, К. Маклеод и М. Маккарти), показало, что носителем генетической информации является ДНК. Этот вывод позже был подтвержден при изучении явления трансдукции (Дж. Ледерберг и М. Зиндер, 1952) — переноса информации от одной бактериальной клетки к другой с помощью фаговой ДНК.
Перечисленные исследования определили повышенный интерес к изучению структуры ДНК, следствием которого явилось создание в 1953 г. модели молекулы ДНК Дж. Уотсоном (амер. биолог) и Ф. Криком (англ. химик). Она была названа двойной спиралью, так как согласно модели построена из двух закрученных в спираль полинуклеотидных цепей. ДНК — полимер, мономерами которого являются нуклеотиды. Каждый нуклеотид состоит из пятиуглеродного сахара дезоксирибозы, остатка фосфорной кислоты, и одного из четырех азотистых оснований (аденин, гуанин, цитозин и тимин). Эта работа сыграла основную роль в дальнейшем развитии генетики и молекулярной биологии.
На основании этой модели был вначале постулирован (Ф. Крик), а затем и доказан экспериментально (М. Месельсон и Ф. Сталь, 1957 г.) полуконсервативный механизм синтеза ДНК, при котором молекула ДНК разделяется на две одиночные цепи, каждая из которых служит матрицей для синтеза дочерней цепи. В основе синтеза лежит принцип комплементарности, определенный ранее Э. Чаргаффом (1945), согласно которому азотистые основания двух цепей ДНК располагаются друг против друга парами, причем аденин соединяется только с тимином (А-Т), а гуанин с цитозином (G-C). Одним из следствий создания модели стала расшифровка генетического кода — принципа записи генетической информации. Над этой проблемой трудились многие научные коллективы в разных странах. Успех пришел к амер. генетику М. Ниренбергу (нобелевский лауреат), в лаборатории которого было расшифровано первое кодовое слово — кодон. Этим словом стал триплет YYY, последовательность из трех нуклеотидов с одним и тем же азотистым основанием — урацилом. В присутствии молекулы иРНК, состоящей из цепочки таких нуклеотидов, синтезировался монотонный белок, содержащий последовательно соединенные остатки одной и той же аминокислоты — фенилаланина. Дальнейшая расшифровка кода была делом техники: используя матрицы с разными сочетаниями оснований в кодонах, ученые составили кодовую таблицу. Были определены все особенности генетического кода: универсальность, триплетность, вырожденность и неперекрываемость. Расшифровку генетического кода по значению для развития науки и практики сравнивают с открытием ядерной энергии в физике.
После расшифровки генетического кода и определения принципа записи генетической информации ученые задумались над тем, каким образом осуществляется перенос информации с ДНК на белок. Исследования этой проблемы закончились полным описанием механизма реализации генетической информации, включающего два этапа: транскрипцию и трансляцию.
После определения химической природы гена и принципа его действия встал вопрос о том, как регулируется работа генов. Впервые он прозвучал в исследованиях французских биохимиков Ф. Жакоба и Ж. Моно (1960), которые разработали схему регуляции группы генов, контролирующих процесс сбраживания лактозы в клетке кишечной палочки. Они ввели понятие бактериального оперона как комплекса, который объединяет все гены (как структурные, так и гены-регуляторы), обслуживающие какое-либо звено метаболизма. Позже правильность их схемы была доказана экспериментально при изучении разнообразных мутаций, затрагивающих различные структурные единицы оперона.
Постепенно вырабатывалась схема механизма регуляции генов эукариот. Этому способствовало установление прерывистой структуры некоторых генов и описание механизма сплайсинга.
Под влиянием прогресса в изучении структуры и функции генов в начале 70-х гг. ХХ в. у генетиков возникла идея манипуляции ими, в первую очередь, путем переноса их из клетки в клетку. Так появилось новое направление генетических исследований — генная инженерия.
Базу для развития этого направления составили эксперименты, в ходе которых были разработаны методы получения отдельных генов. В 1969 г. в лаборатории Дж. Бэквита из хромосомы кишечной палочки с использованием явления трансдукции был выделен лактозный оперон. В 1970 г. коллективом под руководством Г. Корано был впервые осуществлен химический синтез гена. В 1973 г. разработан метод получения фрагментов ДНК — доноров генов — с использованием ферментов рестриктаз, разрезающих молекулу ДНК. И, наконец, был разработан метод получения генов на основе явления обратной транскрипции, открытый в 1975 г. Д. Балтимором и Г. Теминым. Для введения чужеродных генов в клетки на основе плазмид, вирусов, бактериофагов и транспозонов (мобильных генетических элементов) конструировались различные векторы — молекулы-переносчики, которые осуществляли процесс переноса. Комплекс вектора с геном был назван рекомбинантной молекулой. Первая рекомбинантная молекула на основе ДНК фага была сконструирована в 1974 г. (Р. Маррей и Д. Маррей). В 1975 г. были разработаны методы клонирования клеток и фагов со встроенными генами.
Уже в начале 70-х гг. были получены первые результаты экспериментов в области генной инженерии. Так, в клетку кишечной палочки была введена рекомбинантная молекула, содержащая два разных гена устойчивости к антибиотикам (тетрациклину и стрептомицину), после чего клетка приобрела резистентность к обоим препаратам.
Постепенно расширялся набор векторов и вводимых генов и совершенствовалась технология переноса. Это позволило широко использовать методы генной инженерии в промышленных целях (биотехнология), в первую очередь в интересах медицины и сельского хозяйства. Были сконструированы бактерии — продуценты биологически активных веществ. Это позволило наладить в нужных масштабах синтез таких необходимых человеку препаратов, как инсулин, соматостатин, интерферон, триптофан и др. Создано большое количество трансгенных растений, которые стали обладателями ценных свойств (устойчивость к вредителям, засухе, высокое содержание белка и пр.) в результате введения в их геном чужеродных генов.
В 70-х гг. были начаты работы по секвенированию геномов разных объектов, начиная с бактериофагов и кончая человеком.
Особого внимания заслуживает международная генетическая программа “Геном человека”, целью которой являются полная расшифровка генетического кода человека и картирование его хромосом. В перспективе намечается интенсивное развитие новой области медицинской генетики — генотерапии, которое должно способствовать снижению риска проявления вредных генов и тем самым максимальному ограничению генетического груза.
История развития генетики в России
Становление генетики в России произошло во втором десятилетии ХХ в. Создателем первой отечественной школы генетиков был Юрий Александрович Филипченко. В 1916 г. он начал читать в Санкт-Петербургском университете курс лекций “Учение о наследственности и эволюции”, в котором центральное место отвел законам Менделя и исследованиям Т. Моргана. Им был сделан авторизированный перевод книги Моргана “Теория гена”. Научные интересы Ю.А. Филипченко лежали в области наследственности и изменчивости качественных и количественных признаков. Особое внимание он уделял статистическим закономерностям изменчивости. Ю.А. Филипченко написал ряд превосходных книг, среди них учебник “Генетика”, по которому в нашей стране училось несколько поколений биологов.
В этот же период сформировались еще две научные генетические школы: одна в Институте экспериментальной биологии (г. Москва) под руководством Николая Константиновича Кольцова, другая под руководством Николая Ивановича Вавилова начала создаваться в Саратове, где он был избран профессором университета, а окончательно сформировалась в Ленинграде на базе Всесоюзного Института растениеводства (ВИР).
Н.К. Кольцов возглавлял крупный Научно-исследовательский институт экспериментальной биологии в Москве. Он первым высказал идею о макромолекулярной организации носителей наследственности (хромосом) и их самоудвоении как механизме передачи генетической информации. Идеи Н.К. Кольцова оказали сильное влияние на известных ученых того периода, не только биологов, но и физиков, чьи исследования структуры гена привели к развитию молекулярной генетики. Из научной школы Н.К. Кольцова вышли такие крупные генетики, как А.С. Серебровский, Б.Л. Астауров, Н.П. Дубинин, Н.В. Тимофеев-Ресовский, В.В. Сахаров и другие.
Выдающийся генетик и селекционер Н.И. Вавилов завоевал широкое признание своими трудами в области изучения мирового земледелия и растительных ресурсов. Он является автором учения о центрах происхождения и разнообразия культурных растений и учения об иммунитете, а также закона гомологических рядов в наследственной изменчивости. Кроме того, им создана мировая коллекция сельскохозяйственных и технических растений, в том числе знаменитая коллекция сортов пшеницы. Н.И. Вавилов пользовался большим авторитетом не только среди отечественных, но и среди зарубежных ученых. В созданный им в Ленинграде Всесоюзный институт растениеводства (ВИР) съезжались работать ученые со всех стран мира. Признанием заслуг Н.И. Вавилова стало избрание его президентом Международного генетического конгресса, который состоялся в 1937 г. в Эдинбурге. Однако обстоятельства не позволили Н.И. Вавилову присутствовать на этом съезде.
Серьезный вклад в развитие теоретической генетики внесли исследования профессора Московского университета Александра Сергеевича Серебровского и его молодых коллег Н.П. Дубинина, Б.Н. Сидорова, И.И. Агола и других. В 1929 г. ими было сделано открытие явления ступенчатого аллелизма у дрозофилы, которое стало первым шагом к отказу от утвердившегося среди генетиков представления о неделимости гена. Была сформулирована центровая теория строения гена, согласно которой ген состоит из более мелких субъединиц — центров, которые могут мутировать независимо друг от друга. Эти исследования послужили стимулом для развертывания работ по изучению структуры и функции гена, результатом которых стала выработка современной концепции сложной внутренней организации гена. Позже (в 1966 г.) за цикл работ в области теории мутаций Н.П. Дубинин был удостоен Ленинской премии.
К началу 40-х гг. ХХ в. в СССР генетика находилась в состоянии расцвета. Помимо указанных выше, следует отметить работы Б.Л. Астаурова по регулированию пола у тутового шелкопряда генетическими методами; цитогенетические исследования Г.А. Левитского, работы А.А. Сапегина, К.К. Мейстера, А.Р. Жебрака, Н.В. Цицина по генетике и селекции растений; М.Ф. Иванова по генетике и селекции животных; В.В. Сахарова, М.Е. Лобашева, С.М. Гершензона, И.А. Рапопорта по химическому мутагенезу; С.Г. Левита и С.Н. Давиденкова по генетике человека и работы многих других талантливых ученых.
Однако сложившаяся в СССР к началу Второй мировой войны политическая ситуация противостояния капиталистическому миру привела к гонениям на ученых, работавших в области генетики, которая была объявлена идеалистической буржуазной наукой, а ее приверженцы — агентами мирового империализма. Репрессии обрушились на головы многих известных ученых, в том числе Н.И. Вавилова, М.Е. Лобашева, Г.Д. Карпеченко, С.М. Гершензона и многих, многих других. Генетика была отброшена на несколько десятилетий назад. Немалую роль в развале генетической науки сыграл Т.Д. Лысенко. Будучи простым агрономом, он не смог подняться до уровня классической генетики с ее абстрактными представлениями о гене и поэтому просто отрицал законы Менделя, хромосомную теорию наследственности Моргана, учение о мутациях. Свою научную несостоятельность Лысенко прикрывал щедрыми обещаниями быстрого подъема сельского хозяйства с помощью пропагандируемых им методов переделки растений под влиянием условий выращивания, чем заслужил поддержку лично И.В. Сталина. В качестве щита Лысенко использовал работы выдающегося селекционера И.В. Мичурина. В отличие от мировой науки, наша генетика стала называться мичуринской. Такая “честь” привела к тому, что за Мичуриным закрепилась слава приверженца идей Лысенко, которая не покидала ученого даже после краха деятельности последнего. На самом же деле И.В. Мичурин был выдающимся селекционером-практиком, плодоводом, никогда не имевшим отношения к разработке теоретических основ генетической науки.
Отечественная наука окончательно очистилась от “лысенковщины” только к середине 60-х гг. Вышли из “подполья” многие из пострадавших от репрессий ученых, те, кому удалось выжить, в том числе Н.В. Тимофеев-Ресовский, М.Е. Лобашов, В.В. Сахаров и другие. Сохраненные ими традиции и большой потенциал, заложенный в их учениках, способствовали быстрому движению вперед, хотя отставание от мирового уровня, конечно, давало о себе знать. Тем не менее, поднималось новое поколение отечественных генетиков, которым предстояло вывести эту науку на прежний уровень. И снова ряды ученых с мировой известностью пополнились российскими именами: А.Н. Белозерского, В.А. Энгельгардта, С.И. Алиханяна, Р.Б. Хесина, А.С. Спирина, С.В. Шестакова, С.Г. Инге-Вечтомова, Ю.П. Алтухова и многих других.
Однако новые социальные потрясения, вызванные перестройкой, повлекшей отток научных кадров за границу, снова помешали нашей науке обрести соответствующий статус. Остается надеяться, что молодое поколение, опираясь на заложенный предшествующими корифеями фундамент, сможет выполнить эту благородную миссию.
План лекции
Предмет генетики. Сущность явлений наследственности и изменчивости.
Методы генетики.
Краткая история развития генетики.
Генетические исследования в Республике Беларусь
Связь генетики с другими науками.
Значение генетики.
Вопрос. Предмет генетики. Сущность явлений наследственности и изменчивости.
Генетика – это наука о наследственности и изменчивости живых организмов и методах управления ими; это наука, изучающая наследственность и изменчивость признаков живых организмов.
Наследственность – это
1) способность организмов порождать себе подобных;
2) способность организмов передавать (наследовать) свои признаки и качества из поколения в поколение;
3) сохранение определенных вариантов признаков при смене поколений.
Изменчивость – это способность организмов изменяться по признакам тела или отдельных его частей, а также функций.
Изменчивость – это
1) существование признаков в различных формах (вариантах);
2) появление различий между организмами (частями организма или группами организмов) по отдельным признакам.
Основные типы наследования признаков
Прямое наследование , при котором варианты признаков сохраняются в неизменном виде из поколения в поколение.
– при вегетативном размножении растений;
– при самоопылении у растений;
– при размножении чистопородных животных и перекрестном опылении чистосортных растений.
Непрямое наследование – это тип наследования, который наблюдается при половом размножении животных и семенном размножении растений.
Для изучения непрямого наследования необходима гибридизация – скрещивание организмов, различающихся по генотипу.
При непрямом наследовании некоторые варианты признаков проявляются в каждом поколении (такие признаки называются доминантными, «господствующими»), а другие варианты могут временно «исчезать», а затем проявляться в последующих поколениях (такие признаки называются рецессивными, «отступающими»).
Сложные типы наследования признаков заранее предсказать появление новых вариантов признаков очень трудно. В некоторых случаях «внезапно» возникают новые варианты признаков, которых не было ни у родителей, ни у бабушек–дедушек, ни у тетушек–дядюшек. Сложное наследование признаков возможно как на основе влияния условий среды на развитие организма, так и в результате возникновения новых генов или новых комбинаций имеющихся в организме генов.
Вопрос. Методы генетики.
В генетике, как и других науках, используются многочисленные методы при проведении исследований. Для генетики присущи свои специфические методы исследований:
Гибридологический анализ – основной метод, при котором проводят целенаправленное скрещивание родителей с определенными признаками и наблюдают за проявлением этих признаков в поколениях потомков.
Принципы гибридологического анализа:
1. Использование в качестве исходных особей (родителей), форм, не дающих расщепления при скрещивании, т.е. константных форм.
2. Анализ наследования отдельных пар альтернативных признаков, то есть признаков, представленных двумя взаимоисключающими вариантами.
3. Количественный учет форм, выщепляющихся в ходе последовательных скрещиваний и использование математических методов при обработке результатов.
4. Индивидуальный анализ потомства от каждой родительской особи.
5. На основании результатов скрещивания составляется и анализируется схема скрещиваний.
Генеалогический – заключается в анализе родословных и позволяет определить тип наследования (доминантный, рецессивный, аутосомный или сцепленный с полом) признака, а также его моногенность или полигенность. На основе полученных сведений прогнозируют вероятность проявления изучаемого признака в потомстве, что имеет большое значение для предупреждения наследственных заболеваний.
Цитогенетический – изучение хромосом: подсчет их числа, описание структуры, поведения при делении клетки, а также связи изменения структуры хромосом с изменчивостью признаков.
Биохимический – основан на изучении активности ферментных систем. Активность оценивают или по активности самого фермента, или по количеству конечных продуктов реакции, которую контролирует данный фермент. Изучения активности ферментных систем позволяет выявлять генные мутации, которые являются причинами болезней обмена веществ, например, фенилкетонурии, серповидно-клеточной анемии.
Молекулярный – позволяет анализировать фрагменты ДНК, находить и изолировать отдельные гены, устанавливать последовательность нуклеотидов (несут наследственную информацию).
вопрос. Краткая история развития генетики.
Знаменитый врач Древней Греции Гиппократ полагал, что в яйцевой клетке, или в теле матери, должен находиться маленький, но полностью сформированный, преформированный организм. Эти убеждения позже стали называться преформизмом (от лат. preforraatio - предобразование). Спор между преформистами шел только о том, где именно находится этот организм,- в женском или мужском начале.
Противоположные взгляды, согласно которым организм развивается из бесструктурной, гомогенной массы, впервые высказанные еще Аристотелем, в дальнейшем получили развитие и название эпигенеза (от греч. epi - после и genesis - развитие).
Ч. Дарвин впервые поставил биологию на научную основу. Он показал, что в основе эволюции и селекции лежит действие наследственности, изменчивости и отбора. Эти положения стали основой для всего последующего развития генетики.
Первый этап развития науки.
Ознаменовался открытием Г. Менделем (1865) дискретности (делимости) наследственных факторов и разработкой гибридологического метода, изучением наследственности, т. е. правил скрещивания организмов и учета признаков в потомстве.
Значение открытий Г. Менделя оценили после того, как его законы были вновь переоткрыты в 1900 г. тремя биологами независимо друг от друга: де Фризом в Голландии, К. Корренсом в Германии и Э. Чермаком в Австрии.
В 1901 -1903 гг. Хуго де Фриз выдвинул мутационную теорию изменчивости, которая сыграла большую роль в дальнейшем развитии генетики.
Важное значение имели работы датского ботаника Вильгельма Людвига Иоганнсена, который изучал закономерности наследования на чистых линиях фасоли. Он сформулировал также понятие «популяция» (группа организмов одного вида, обитающих и размножающихся на ограниченной территории), предложил называть менделевские “наследственные факторы” термином «ген», дал определения понятий “генотип” и “фенотип”.
Второй этап
Характеризуется переходом к изучению явлений наследственности на клеточном уровне (цитогенетика). Т. Бовери (1902-1907 гг.), У. Сэттон и Э. Уильсон (1902-1907 гг.) установили взаимосвязь между менделевскими законами наследования и распределением хромосом в процессе клеточного деления (митоз) и созревания половых клеток (мейоз).
Решающее значение в обосновании хромосомной теории наследственности имели исследования, проведенные на мушках дрозофилах американским генетиком Т. Г. Морганом и его сотрудниками (1910-1911 гг.).
Морган установил также закономерности наследования признаков, сцепленных с полом.
Следующим шагом было установление химической природы хромосомных генов. Советский генетик Н.К. Кольцов одним из первых развил представление об их макромолекулярной природе (1927 г.), а Н.В. Тимофеев-Ресовский с соавторами в середине 30-х гг. 20 в. вычислил примерный объем гена.
Впервые в 1925 г. советские микробиологи Г.А. Надсон и Г.С. Филиппов показали, что после облучения дрожжевых клеток ионизирующим излучением возникают разнообразные радиорасы, свойства которых воспроизводятся в потомстве. В 1927 г. Мёллер (Н.J. Muller) в точных опытах на дрозофилах с учетом дозы облучения установил возникновение новых наследственных мутаций. Позже И.А. Рапопорт и Ауэрбах (Ch. Auerbach) открыли явление мутагенеза под влиянием химических веществ.
Третий этап
Отражает достижения молекулярной биологии и связан с использованием методов и принципов точных наук - физики, химии, математики, биофизики и др.- в изучении явлений жизни на уровне молекул. Объектами генетических исследований стали грибы, бактерии, вирусы.
На этом этапе были изучены взаимоотношения между генами и ферментами и сформулирована теория “один ген - один фермент” (Дж. Бидл и Э. Тейтем, Дж. Ледерберг, 1940): каждый ген контролирует синтез одного фермента; фермент в свою очередь контролирует одну реакцию из целого ряда биохимических превращений, лежащих в основе проявления внешнего или внутреннего признака организма.
В 1953 г. Ф. Крик и Дж. Уотсон, опираясь на результаты опытов генетиков и биохимиков, на данные рентгеноструктурного анализа, создали структурную модель ДНК в форме двойной спирали. Предложенная ими модель ДНК хорошо согласуется с биологической функцией этого соединения: способностью к самоудвоению генетического материала и устойчивому сохранению его в поколениях - от клетки к клетке.
В последнее десятилетие возникло новое направление в молекулярной генетике - генная инженерия - система приемов, позволяющих биологу конструировать искусственные генетические системы.
Федеральное агентство по образованию Российской Федерации
Государственное образовательное учреждение высшего профессионального образования
«Южно-Уральский государственный университет»
Факультет «Экономика и управление»
Кафедра «Экономика, управление и инвестиции»
История развития генетики. Вклад русских ученых
РЕФЕРАТ
по дисциплине «Концепции современного естествознания»
Проверил
О.М. Баева
студент группы ЭиУ-232
А.И. Кулешова
________________________2010г.
Реферат защищен
с оценкой
_____________________________
________________________2010г.
ВВЕДЕНИЕ
Генетика - наука о наследственности и её изменчивости – получила развитие в начале XX в., после того как исследователи обратили внимание на законы Г. Менделя, открытые в 1865 г., но остававшиеся без внимания в течение 35 лет. За короткий срок генетика выросла в разветвленную биологическую науку с широким кругом экспериментальных методов и направлений. Название генетика было предложено английским ученым У. Бэтсоном в 1906 г. Исследователями классического периода развития генетики были выяснены основные закономерности наследования и доказано, что наследственные факторы (гены) сосредоточены в хромосомах. Дальнейший прогресс в изучении закономерностей хранения и реализации генетической информации сдерживался по двум причинам. Во-первых, из-за слишком объемных экспериментов, связанных с более глубоким изучением генов, во-вторых, ввиду невозможности понять работу генов без углубленного исследования превращения молекул, вовлеченных в генетические процессы. Переход к генетическим исследованиям микроорганизмов, позволивший избегать многих трудностей, был вполне закономерен. Такой переход осуществился в 50-х годах. В 1941 г. Дж. Бидл и Э. Тейтум опубликовал короткую статью "Генетический контроль биохимических реакций у Neurospora ", в которой сообщили о первых генетических экспериментах на микроорганизмах.
В последние годы эти исследования получили широкий размах и проводятся на самых различных биологических объектах.
Задачей данного реферата является отражение наиболее важных открытий, сделанных русскими учеными в области генетики, их анализ и определение их значимости для науки.
Для раскрытия темы были взяты как научные труды, так и современные интернет-ресурсы, что должно дать проверенные данные и современную точку зрения на них.
1 РАЗВИТИЕ ГЕНЕТИКИ В РОССИИ
Если не считать опытов по гибридизации растений в XVIII в., первые работы по генетике в России были начаты в начале XX в. как на опытных сельскохозяйственных станциях, так и в среде университетских биологов, преимущественно тех, кто занимался экспериментальной ботаникой и зоологией.
После революции и гражданской войны 1917-1922 гг. началось стремительное организационное развитие науки. К концу 1930-х годов в СССР была создана обширная сеть научно-исследовательских институтов и опытных станций (как в Академии наук СССР, так и во Всесоюзной академии сельскохозяйственных наук имени Ленина (ВАСХНИЛ)), а также вузовских кафедр генетики. Признанными лидерами направления были Н. И. Вавилов, Н. К. Кольцов, А. С. Серебровский, С. С. Четвериков и др. В СССР издавали переводы трудов иностранных генетиков, в том числе Т. Х. Моргана, Г. Мёллера, ряд генетиков участвовали в международных программах научного обмена. Американский генетик Г. Мёллер работал в СССР (1934-1937), советские генетики работали за границей. Н. В. Тимофеев-Ресовский - в Германии (с 1925 г.), Ф. Г. Добржанский - в США (с 1927 г.).
В 1930-е гг. в рядах генетиков и селекционеров наметился раскол, связанный с энергичной деятельностью Т. Д. Лысенко и И. И. Презента. По инициативе генетиков был проведён ряд дискуссий (наиболее крупные - в 1936 и 1939 г.), направленных на борьбу с подходом Лысенко.
На рубеже 1930-1940-х гг. в ходе так называемого Большого террора большинство сотрудников аппарата ЦК ВКП (б), курировавших генетику, и ряд видных генетиков были арестованы, многие расстреляны или погибли в тюрьмах (в том числе, Н. И. Вавилов). После войны дебаты возобновились с новой силой. Генетики, опираясь на авторитет международного научного сообщества, снова попытались склонить чашу весов в свою сторону, однако с началом холодной войны ситуация значительно изменилась. В 1948 году на августовской сессии ВАСХНИЛ Т. Д. Лысенко, пользуясь поддержкой И. В. Сталина, объявил генетику лженаукой. Лысенко воспользовался некомпетентностью партийного руководства наукой, «пообещав партии» быстрое создание новых высокопродуктивных сортов зерна («ветвистая пшеница») и др. С этого момента начался период гонений на генетику, который получил название лысенковщины и продолжался вплоть до снятия Н. С. Хрущева с поста генерального секретаря ЦК КПСС в 1964 г.
Лично Т. Д. Лысенко и его сторонники получили контроль над институтами отделения биологии АН СССР, ВАСХНИЛ и вузовскими кафедрами. Были изданы новые учебники для школ и вузов, написанные с позиций «Мичуринской биологии». Генетики вынуждены были оставить научную деятельность или радикально изменить профиль работы. Некоторым удалось продолжить исследования по генетике в рамках программ по изучению радиационной и химической опасности за пределами организаций, подконтрольных Т. Д. Лысенко и его сторонникам.
После открытия и расшифровки структуры ДНК, физической базы генов (1953 г.), с середины 1960-х г. началось восстановление генетики. Министр просвещения РСФСР В. Н. Столетов инициировал широкую дискуссию между лысенковцами и генетиками, в результате было опубликовано много новых работ по генетике. В 1963 г. вышел в свет университетский учебник М. Е. Лобашева «Генетика», выдержавший впоследствии несколько изданий. Вскоре появился и новый школьный учебник Общая биология под редакцией Ю. И. Полянского, используемый, наряду с другими, и по сей день.
Вывод по разделу один
Развитие генетики в России шло сложным путем, претерпевая гонения со стороны властных структур, что значительно тормозило процесс развития данной науки.
2 НИКОЛАЙ ИВАНОВИЧ ВАВИЛОВ И ЕГО ВКЛАД В ГЕНЕТИКУ
Николай Иванович Вавилов (13 (25) ноября 1887, Москва, Российская империя - 26 января 1943, Саратов, РСФСР, СССР) - российский и советский учёный-генетик, ботаник, селекционер, географ, академик АН СССР, АН УССР и ВАСХНИЛ. Президент (1929-1935), вице-президент (1935-1940) ВАСХНИЛ, президент Всесоюзного географического общества (1931-1940), основатель (1920) и бессменный до момента ареста директор Всесоюзного института растениеводства (1930-1940), директор Института генетики АН СССР (1930-1940), член Экспедиционной комиссии АН СССР, член коллегии Наркомзема СССР, член президиума Всесоюзной ассоциации востоковедения. В 1926-1935 годах член Центрального исполнительного комитета СССР, в 1927-1929 - член Всероссийского Центрального Исполнительного Комитета.
Организатор и участник ботанико-агрономических экспедиций, охвативших большинство континентов (кроме Австралии и Антарктиды), в ходе которых выявил древние очаги формообразования культурных растений. Создал учение о мировых центрах происхождения культурных растений. Обосновал учение об иммунитете растений, открыл закон гомологических рядов в наследственной изменчивости организмов. Внёс существенный вклад в разработку учения о биологическом виде. Под руководством Вавилова была создана крупнейшая в мире коллекция семян культурных растений. Он заложил основы системы государственных испытаний сортов полевых культур. Сформулировал принципы деятельности главного научного центра страны по аграрным наукам, создал сеть научных учреждений в этой области.
2.1 Учение об иммунитете растений
Вавилов подразделял иммунитет растений на структурный (механический) и химический. Механический иммунитет растений обусловлен морфологическими особенностями растения-хозяина, в частности, наличием защитных приспособлений, которые препятствуют проникновению патогенов в тело растений. Химический иммунитет зависит от химических особенностей растений.
2.2 Закон гомологических рядов в наследственной изменчивости
В работе «Закон гомологических рядов в наследственной изменчивости» Вавиловым было введено понятие «Гомологические ряды в наследственной изменчивости». Понятие было введено при исследовании параллелизмов в явлениях наследственной изменчивости по аналогии с гомологическими рядами органических соединений.
Суть явления состоит в том, что при изучении наследственной изменчивости у близких групп растений были обнаружены сходные аллельные формы, которые повторялись у разных видов (например, узлы соломины злаков с антоциановой окраской или без неё, колосья с остью или без неё и т. п.). Наличие такой повторяемости давало возможность предсказывать наличие ещё не обнаруженных аллелей, важных с точки зрения селекционной работы. Поиск растений с такими аллелями проводился в экспедициях в предполагаемые центры происхождения культурных растений. Следует помнить, что в те годы искусственная индукция мутагенеза химическими веществами или воздействием ионизирующих излучений ещё не была известна, и поиск необходимых аллелей приходилось производить в природных популяциях.
Первая (1920) формулировка закона включала в себя две закономерности:
Первая закономерность, которая бросается в глаза при детальном изучении форм у каких-либо линнеонов растений, принадлежащих к одному и тому же роду, - это тождество рядов морфологических и физиологических свойств, характеризующих разновидности и расы у близких генетических линнеонов, параллелизм рядов видовой генотипической изменчивости. Чем ближе генетически виды, тем резче и точнее проявляется тождество рядов морфологических и физиологических признаков.
2-я закономерность в полиморфизме, вытекающая по существу из первой, состоит в том, что не только генетически близкие виды, но и роды проявляют тождества в рядах генотипической изменчивости.
В 1923 году Вавилов включил обсуждение закона в работу «Новейшие успехи в области теории селекции», в которой показал, что благодаря закономерности проявления сортовых различий у видов и родов «можно определённо предвидеть и находить соответствующие формы у изучаемого растения». Действительно, на основе закона гомологических рядов Вавилов и его сотрудники сотни раз предугадывали существование тех или иных форм, а затем и обнаруживали их. Вавилов отмечал, что «общие ряды изменчивости свойственны иногда и очень отдалённым, генетически не связанным семействам». Вавилов допускал, что ряды параллельной изменчивости не обязательно будут полными и будут лишены некоторых звеньев в результате действия естественного отбора, летальных сочетаний генов и вымирания видов. Однако, «несмотря на огромную роль естественного отбора и вымирание многих связующих звеньев, … не представляет затруднений проследить сходство в наследственной изменчивости у близких видов».
Хотя закон был открыт в результате изучения фенотипической изменчивости, Вавилов распространил его действие и на генотипическую изменчивость: «Исходя из поразительного сходства в фенотипической изменчивости видов в пределах одного и того же рода или близких родов, обусловленного единством эволюционного процесса, можно предполагать наличие у них множества общих генов наряду со спецификой видов и родов».
Вавилов считал, что закон справедлив не только по отношению к морфологическим признакам, предвидя, что уже установленные ряды «не только будут пополняться недостающими звеньями в соответствующих клетках, но и будут развиваться, в особенности в отношении физиологических, анатомических и биохимических признаков». В частности, Вавилов отметил, что близкие виды растений характеризуются «сходством химического состава, выработкой близких или одних и тех же специфических химических соединений». Как было показано Вавиловым, внутривидовая изменчивость химического состава (например, эфирных масел и алкалоидов) касается главным образом количественных соотношений при постоянстве качественного состава, тогда как в пределах рода химический состав отдельных видов отличается и количественно, и качественно. При этом в пределах рода «отдельные виды обычно характеризуются теоретически предусматриваемыми химиками изомерами или производными и обычно связаны между собой взаимными переходами». Параллелизм изменчивости характеризует близкие роды с такой определённостью, что «им можно пользоваться в поисках соответствующих химических компонентов», а также «получать синтетически в пределах данного рода при помощи скрещивания химические вещества определённого качества».
Вавилов выяснил, что закон проявляется не только в пределах родственных групп; параллелизм изменчивости был обнаружен «в разных семействах, генетически не связанных, даже в разных классах», но в отдалённых семействах параллелизм не всегда носит гомологичный характер. «Сходные органы и само их сходство являются в данном случае не гомологичными, а только аналогичными».
Закон гомологических рядов не снимал всех трудностей, поскольку было ясно, что одинаковые изменения фенотипических признаков могут быть обусловлены разными генами, а существовавший в те годы уровень знаний не позволял непосредственно связывать признак с определённым геном. В отношении видов и родов Вавилов отмечал, что «мы имеем дело пока в основном не с генами, о которых мы знаем очень мало, а с признаками в условиях определённой среды», и на этом основании предпочитал говорить о гомологичных признаках. «В случае параллелизма отдалённых семейств, классов, конечно, не может быть и речи о тождественных генах даже для сходных внешне признаков».
Несмотря на то, что первоначально закон был сформулирован на основе изучения преимущественно культурных растений, позднее, рассмотрев явление изменчивости у грибов, водорослей и животных, Вавилов пришёл к выводу, что закон носит всеобщий характер и проявляется «не только у высших, но и у низших растений, равно как и у животных».
Прогресс генетики оказал значительное влияние на дальнейшее развитие формулировки закона. В 1936 году Вавилов назвал первую формулировку излишне категорической: «Таково было тогда состояние генетики…». Было принято думать, что «гены идентичны у близких видов», биологи «представляли ген более стабильным, чем в настоящее время». Позже было установлено, что и «близкие виды могут при наличии сходных внешне признаков характеризоваться многими различными генами». Вавилов отмечал, что в 1920 году уделил «мало…внимания роли отбора», сосредоточив основное внимание на закономерностях изменчивости. Это замечание отнюдь не означало забвения теории эволюции, ибо, как подчёркивал сам Вавилов, уже в 1920 году его закон «прежде всего представлял формулу точных фактов, основанных всецело на эволюционном учении».
Вавилов рассматривал сформулированный им закон как вклад в популярные в то время представления о закономерном характере изменчивости, лежащей в основе эволюционного процесса (например, теория номогенеза Л. С. Берга). Он полагал, что закономерно повторяющиеся в разных группах наследственные вариации лежат в основе эволюционных параллелизмов и явления мимикрии.
Вывод по разделу два
Являясь одним из первых генетиков России, Вавилов внес существенный вклад в развитие данной науки, положив начало изучению генетической природы растений.
3 НИКОЛАЙ КОНСТАНТИНОВИЧ КОЛЬЦОВ
Николай Константинович Кольцов (3 (15) июля 1872, Москва - 2 декабря 1940 года, Ленинград) - выдающийся русский биолог, автор идеи матричного синтеза.
Начиная свою работу в период расцвета описательной биологии и первых шагов экспериментальной биологии, Кольцов тонко чувствовал тенденции развития биологии и рано осознал значение экспериментального метода. Он проповедовал необходимость экспериментального подхода во всех областях биологии и предсказал его использование даже в эволюционном учении (не противопоставляя экспериментальные методы описательным). Речь шла не о простом биологическом эксперименте, а об использовании методов физики и химии. Кольцов не раз подчеркивал огромное значение для биологии открытия новых форм лучистой энергии, в частности рентгеновских и космических лучей, писал о применении радиоактивных веществ. Чтобы изучить организм в целом, надо использовать все современные знания в области физической и коллоидной химии, необходимо изучать внутри клетки мономолекулярные слои и их роль в разнообразных превращениях веществ. "Биологи ждут, когда эти методы (рентгеноструктурного анализа) будут усовершенствованы настолько, что можно будет при их помощи изучить кристаллическую структуру внутриклеточных скелетных, твердых структур белкового и иного характера". Эта мысль явилась пророческой и реально осуществилась в открытии методом рентгеноструктурного анализа строения молекулы ДНК. Пророческой оказалась и другая идея Кольцова, в которой он также шел от биологии к химии. Исходя из развиваемого им представления, что каждая сложная биологическая молекула возникает из подобной ей уже существующей молекулы, он предсказал, что химики пойдут по пути создания новых молекул в растворах, содержащих необходимые составные части сложных молекул, путем внесения в них затравок готовых молекул той же структуры. Он писал: "Я думаю, что только таким способом удастся синтезировать in vitro белки, и при том не какие-нибудь, а определенные, т. е. синтез которых заранее намечается".
В поле зрения Н.К.Кольцова постоянно находились вопросы генетики. Еще в 1921 г. им была опубликована экспериментальная работа "Генетический анализ окраски у морских свинок". Проводились генетические исследования на дрозофиле. В этих работах ученый видел установление важнейшей связи между генетикой и эволюционным учением. Позднее начались работы по химическому мутагенезу.
Н.К.Кольцов глубоко понимал значение генетики для практики животноводства. В 1918 г. им была организована Аниковская генетическая станция, специализирующаяся по генетике сельскохозяйственных животных. Несколько позже была организована в Тульской области другая станция по птицеводству. В начале 1920 г. обе станции слились в одну. В 1925 г. станция получила название Центральной станции по генетике сельскохозяйственных животных, директором которой в разные годы был Кольцов и его ученики.
Вывод по разделу три
Н.К. Кольцов предугадал дальнейшее развитие генетики, указав таким образом своим последователям путь. Именно благодаря ему и его идеям было сделано множество открытий, таких как химический мутагенез. Еще одна заслуга Кольцова состоит в том, что он привлек для работы на станции много талантливых людей известных впоследствии как создатели целых направлений в генетике и селекции отдельных видов сельскохозяйственных животных.
4 ИСКУССТВЕННОЕ ПОЛУЧЕНИЕ МУТАЦИЙ
Крупнейшим достижением экспериментальной генетики было обнаружение возможности искусственно вызывать мутации при помощи разнообразных физических и химических агентов. Немалый вклад в открытие этой возможности внесли русские ученые.
4.1 Вклад Г.А. Надсона и его учеников
Одними из первых опыты в данной области проделали русские ученые Георгий Адамович Надсон и его молодой сотрудник Григорий Семенович Филиппов, работавшие в Институте рентгенологии и радиологии.
Они получили мутации у дрожжей и низших грибов под действием радия и рентгенных лучей. В результате экспериментов вырастали колонии, отличавшиеся величиной, формой, окраской, дрожжевые клетки изменяли свои биохимические свойства. Таким образом была открыта способность радиации вызывать наследственные изменения.
Также Г.А. Надсон совместно с Э.Я. Рохлиной опубликовал ряд работ, в которых впервые поднимался вопрос о практическом использовании экспериментально полученных мутантов.
Кроме дрожжей и низших грибов, в лабораториях Г.А.Надсона изучалось генетическое действие излучений на бактерии. Было заложено новое направление в науке - популяционная генетика микроорганизмов. Значительное внимание Г.А.Надсон уделял также изучению и анализу роли внутренних и внешних факторов в процессе экспериментального мутагенеза, например, почему разные виды микроорганизмов по-разному реагируют на облучение и как интенсивность и доза облучения влияют на индукцию мутаций. Еще одно направление работ школы Г.А.Надсона - химический мутагенез. Его учениками в 1928 г. были получены данные о возникновении наследственных изменений у дрожжей под действием хлороформа, в 1939 г. - под действием каменноугольной смолы и цианистого калия.
4.2 Вклад Н.В. Тимофеева-Ресовского
В середине 30-х годов была сформулирована теория, описывающая кинетические зависимости ни активирующего и мутагенного эффекта ионизирующих излучений - так называемая "теория мишени". Важнейшие эксперименты, ставшие основой этой теории, были проведены в период 1931 - 1937 гг. несколькими исследователями, среди которых был и Николай Владимирович Тимофеев-Ресовский, ставший одним из основателей количественной биофизики ионизирующих излучений.
Тимофеев-Ресовский развивал идеи Н. Кольцова, предполагавшего, что молекулярные наследственные структуры образуются посредством матричного синтеза. Он вел исследования по биофизическому анализу мутационного процесса, впоследствии приведшие к формированию молекулярной биологии как новой синтетической дисциплины. Тимофеев-Ресовский показал, что мутационные изменения затрагивают относительно ограниченную группу атомов в хромосоме. Это открытие впервые переводило мутационный процесс на молекулярный уровень понимания.
Также Николай Владимирович считается одним из основоположников радиобиологии. Ему удалось установить, как влияет доза излучения на интенсивность мутационного процесса. Он обнаружил явление радиостимуляции малыми дозами и провел анализ первичных пусковых механизмов возникновения мутаций под влиянием излучений.
Этот исследователь первым указал на то, что помимо прямых последствий воздействия ионизирующего излучения (т. е. злокачественных новообразований, ожогов, лучевой болезни) существует серьезная опасность возникновения вредных мутаций и накопления их в популяциях.
Одной из важнейших составных частей количественной теории мутационного процесса стали исследования русского ученого по вероятностям возникновения прямых и обратных мутаций.
В 1934 году Тимофеев-Ресовский провел ряд блестящих экспериментов, впервые показавших, что сочетание нескольких рецессивных мутаций, каждая из которых порознь снижает жизнеспособность, может привести к повышению жизнеспособности особей - носителей этих комбинаций. Данные исследования позволили в полной мере понять эволюционное значение явлений рецессивности и доминантности.
Совместно с М. Дельбрюком (впоследствии лауреатом Нобелевской премии) Николай Владимирович Тимофеев-Ресовский провел работы по моделированию структуры генов. В тот же период в соавторстве с физиком Р. Ромпе он открыл и описал «принцип усилителя» в биологии, ставший одним из общих важнейших принципов современной теоретической биологии. Согласно этому принципу, единичное изменение способно изменить свойства целой особи и привести в действие силы, на несколько порядков большие по затрачиваемой энергии.
4.3 Химический мутагенез
Первым, кто обратил внимание на мутации, вызванные химическими веществами, был Н.К.Кольцов, предложивший своему последователю, В.В. Сахарову, продолжить исследования в данной области.
В.В. Сахаров, работая по замыслу Кольцова, использовал 10% раствор йода. По анализу видимых мутаций, сцепленных с полом, в потомстве второго поколения он нашел передаваемые по наследству новые мутации Notch и "prune" .
Впервые в мире в 1938 г. В.В. Сахаров сформулировал идею "о специфическом воздействии мутационных факторов", показал различие природы мутаций, возникших спонтанно, и индуцированных физическими и химическими мутагенами.
В наше время сформулированное В.В. Сахаровым положение о том, что специфичность мутагенеза обусловлена как структурой воздействующего фактора, так и особенностями организма, не утратило своего значения и является одним из важнейших обобщений генетики. Цикл работ по изучению мутационного процесса привел к обнаружению роли внутренних факторов (старение, инбридинг и гибридизация) в этом процессе.
Одним из первых В.В. Сахаров поднял вопрос о мутационном последействии колхицина и других мутагенов на растения, что в наши дни обрело ценность, став первоисточником для экологов и исследователей малых доз биологически активных веществ.
Другой исследователь М.Е.Лобашев первым связал процесс возникновения мутаций с репарацией поврежденных генетических структур.
Первые мутагены, открытые Сахаровым, Лобашевым и их сотрудниками, обладали низкой эффективностью, поэтому они не могли заинтересовать практиков.
Новый этап изучения роли химических факторов в процессе мутации был открыт И.А. Рапопортом (1943,46,47) указавшим на мощное мутагенное действие некоторых химических веществ. Он так же организовал широкие работы по использованию химических мутагенов в практике селекции сельскохозяйственных растений.
Вывод по разделу четыре
Русские ученые одними из первых во всем мире начали проводить эксперименты по искусственному получению мутаций, что позволило им сделать выводы о факторах, оказывающих влияние на изменчивость организмов, возможности практического использования новых организмов и возможных последствиях мутаций.
5 ПРОБЛЕМА ДРОБИМОСТИ ГЕНА
К началу 30-х годов XX в. сложились основы теории гена. Уже первые достижения гибридологического анализа подняли проблему дискретности наследственного материала. Считалось, что ген отвечает за развитие одного признака и передается при скрещиваниях как неделимое целое. Открытие мутации и кроссинговера (нарушения сцепления генов в результате обмена участками между хромосомами) подтверждали неделимость генов. В результате обобщения всех данных определение гена получило следующую формулировку: ген - это элементарная единица наследственности, характеризующаяся вполне определенной функцией, мутирующая во время кроссинговера как целое. Иначе говоря, ген – единица генетической функции, мутации и кроссинговера.
В 1928 г. в лаборатории А.С. Серебровского в Биологическом институте им. К.А. Тимирязева Н.П. Дубинин начал исследовать действие рентгеновых лучей на дрозофил и обнаружил необычную мутацию. Образование щетинок на теле мухи контролируется особым геном scute . Мутация гена scute , впервые обнаруженная американским генетиком Пейном (1920), не раз возникала в экспериментах, и при её появлении подавлялось развитие девяти щетинок. Выявленная Дубининым мутация, подавляла развитие всего четырех щетинок. После дальнейших экспериментов стало ясно, что ген не является неделимой генетической структурой, представляет собой область хромосомы, отдельные участки которой могут мутировать независимо друг от друга. Это явление Серебровским ступенчатым аллеломорфизмом.
Одним из крупных достоинств работ по изучению ступенчатых аллеломорфов был количественный метод учета мутантов. Разработав систему, позволяющую количественно оценивать результат каждой мутации, Серебровский, Дубинин и другие авторы тогда же раскрыли явление дополнения одного мутантного гена другим. Это явление было впоследствии переоткрыто на микроорганизмах и получило название комплементации.
Показав мутационную дробимость гена, Серебровский и другие сотрудники его лаборатории, тем не менее, долгое время не могли подтвердить дробимость гена при помощи кроссинговера. Чтобы обнаружить разрыв гена, требовалось проверить огромное число мух. Организовать такой эксперимент удалось только в 1938 г., когда Дубинин, Н.Н. Соколов и Г.Г. Тиняков смогли разорвать ген scute и проверить свой результат цитологически на гигантских хромосомах слюнных желез дрозофилы.
Вывод по разделу пять
Хотя окончательное решение вопроса, делим ли ген не только мутационно, но и механически, и не было достигнуто в работах русских ученых, они внесли существенный вклад в доказательство делимости гена, что повлекло за собой переход к генетическим исследованиям на микроорганизмах, а значит и переход генетики на молекулярный уровень исследований.
6 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА
Выяснение основной функции гена как хранителя информации о строении определенной полипептидной цепи поставило перед молекулярной генетикой вопрос: каким образом осуществляется перенос информации от генетических структур (ДНК) к морфологическим структурам, другими словами, каким образом записана генетическая информация и как она реализуется в клетке.
Согласно модели Уотсона - Крика, генетическую информацию в ДНК несет
последовательность расположения оснований. Таким образом, в ДНК заключены четыре элемента генетической информации. В тоже время в белках было обнаружено 20 основных аминокислот. Необходимо было выяснить, как язык четырехбуквенной записи в ДНК может быть переведен на язык двадцати буквенной записи в беках. Решающий вклад в разработку этого механизма был внесен Г. Гамовым. Он предположил, что для кодирования одной аминокислоты используется сочетание из трех нуклеотидов ДНК (нуклеотидом называют соединение, состоящее из сахара {дизоксорибоза}, фосфата и основания и образующее элементарный мономер ДНК). Эта элементарная единица наследственного материала, кодирующая одну аминокислоту, получила название кодона.
Вывод по разделу шесть
На данном этапе один из русских ученых сделал важный шаг к разгадке генетического кода.
ЗАКЛЮЧЕНИЕ
По материалам данной работы можно сделать вывод о том, что вклад русских ученых в развитие генетики как науки достаточно велик, однако недооценен. Объясняется это во многом неизвестностью русских научных журналов, публиковавших работы ученых, за рубежом.
Однако значимость открытий, сделанных русскими учеными, остается бесспорной.
Достаточно сложно подробно описать работу, проделанную русскими генетиками, поскольку их эксперименты сложны и многообразны. В рамках данной работы отражены лишь ключевые открытия, определившие дальнейшее развитие науки.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Новая Российская энциклопедия: в 12 т. / Редкол.: А. Д. Некипелов, В. И. Данилов-Данильян и др. - М.: Энциклопедия, 2007. Т. 3: Бруней - Винча. - 480 с.
2. Биология – История развития генетики. - http://www.5ka.ru/10/24545/1.html
3. Википедия – Генетика. - http://ru.wikipedia.org/wiki/Генетика
4. Википедия – Вавилов Николай Иванович. - http://ru.wikipedia.org/wiki/Вавилов,_Николай_Иванович
5. Николай Кольцов. - http://idbras.comcor.ru/personal/kolzov.htm
6. Владимир Сахаров. - http://idbras.comcor.ru/personal/SakharovVVr.html
7. Надсон Георгий Адамович. - http://persona.rin.ru/view/f/0/23965/nadson-georgij-adamovich
8. Великие ученые ХХ века – Николай Владимирович Тимофеев-Ресовский. - http://iomn.net/?p=79
 Оригинальные, красивые и веселые поздравления женщине с юбилеем Веселые поздравлялки с днем рождения
Оригинальные, красивые и веселые поздравления женщине с юбилеем Веселые поздравлялки с днем рождения Несколько советов о том, как ускорить высыхание обычного лака на ногтях Как сделать чтобы ногти быстро высохли
Несколько советов о том, как ускорить высыхание обычного лака на ногтях Как сделать чтобы ногти быстро высохли Как его видит правительство
Как его видит правительство