Metode za preučevanje človeške dednosti. Genealoška metoda za proučevanje človeške genetike Primer genealoške metode sestave rodovnika
Metode, ki se pogosto uporabljajo pri preučevanju človeške genetike, vključujejo genealoške, populacijsko-statistične, dvojčke, dermatoglifne, citogenetske, biokemične metode genetike somatskih celic.
genealoška metoda
Osnova te metode je sestavljanje in analiza rodovnikov. Ta metoda se od antičnih časov do danes pogosto uporablja v konjereji, selekciji dragocenih linij goveda in prašičev, pri pridobivanju čistokrvnih psov, pa tudi pri vzreji novih pasem kožuharjev. Človeška rodoslovja so bila sestavljena skozi več stoletij v zvezi s vladajočimi družinami v Evropi in Aziji.
Kot metodo proučevanja človeške genetike se je genealoška metoda začela uporabljati šele v začetku 20. stoletja, ko je postalo jasno, da je mogoče nadomeščati analizo rodovnikov, v kateri je mogoče nadomestiti prenos neke lastnosti (bolezni) iz roda v rod. s hibridno metodo, ki je za človeka dejansko neuporabna.
Pri sestavljanju rodovnikov je vir oseba - proband, katerega rodovnik se preučuje. Običajno je to bodisi bolnik ali nosilec določene lastnosti, katere dedovanje je treba preučiti. Pri sestavljanju rodoslovnih tabel se uporabljajo simboli, ki jih je leta 1931 predlagal G. Yust (slika 7.24). Generacije so označene z rimskimi številkami, posamezniki v dani generaciji so označeni z arabskimi številkami.
S pomočjo genealoške metode je mogoče ugotoviti dedno pogojenost proučevane lastnosti, pa tudi tip
riž. 7.24. konvencije pri sestavljanju rodovnikov (po G. Yustu) njegovega dedovanja (avtosomno dominantno, avtosomno recesivno, X-vezano dominantno ali recesivno, Y-vezano). Pri analizi rodovnikov za več lastnosti je mogoče razkriti povezano naravo njihovega dedovanja, ki se uporablja pri sestavljanju kromosomskih kart. Ta metoda omogoča preučevanje intenzivnosti procesa mutacije in oceno ekspresivnosti in penetracije alela. Široko se uporablja v medicinskem genetskem svetovanju za napovedovanje potomcev. Vendar je treba opozoriti, da postane genealoška analiza veliko bolj zapletena, ko imajo družine malo otrok.
Za avtosomni tip dedovanja je na splošno značilna enaka verjetnost pojava te lastnosti pri moških in ženskah. To je posledica enakega dvojnega odmerka genov, ki se nahajajo v avtosomih vseh predstavnikov vrste in so jih prejeli od obeh staršev, in odvisnosti lastnosti v razvoju od narave interakcije alelnih genov.
Kadar lastnost prevladuje pri potomcih starševskega para, kjer je vsaj eden od staršev njen nosilec, se kaže z večjo ali manjšo verjetnostjo, odvisno od genetske konstitucije staršev (slika 7.25).
Če analiziramo lastnost, ki ne vpliva na sposobnost preživetja organizma, so lahko nosilci prevladujoče lastnosti tako homo- kot heterozigoti. Kdaj prevladujoče dedovanje nekatere patološke lastnosti (bolezni) homozigoti praviloma niso sposobni preživeti, nosilci te lastnosti pa so heterozigoti.
Tako se pri avtosomno dominantnem dedovanju lahko lastnost enako pojavlja pri moških in ženskah in jo je mogoče zaslediti z zadostnim številom potomcev v vsaki generaciji vzdolž vertikale. Pri analizi rodovnikov se je treba spomniti možnosti nepopolnega prodora prevladujočega alela zaradi interakcije genov ali okoljskih dejavnikov. Indeks penetracije lahko izračunamo kot razmerje med dejanskim številom nosilcev lastnosti in številom pričakovanih nosilcev te lastnosti v dani družini. Ne smemo pozabiti, da se nekatere bolezni ne pojavijo takoj od rojstva.

riž. 7.25. Verjetnost, da bi imeli potomce dominantna lastnost iz različnih zakonskih parov (/ - III)
otrok. Številne bolezni, podedovane po dominantnem tipu, se razvijejo šele v določeni starosti. Tako se Huntingtonova koreja klinično manifestira do 35-40 let, pozno se pokaže tudi policistična ledvična bolezen. Zato se pri napovedovanju takšnih bolezni ne upoštevajo bratje in sestre, ki niso dosegli kritične starosti.
Prvi opis rodovnika z avtosomno prevladujočim tipom dedovanja anomalije pri ljudeh je bil podan leta 1905. Sledi prenos v več generacijah. brahidaktilija(short-fingered™). Na sl. 7.26 prikazuje rodovnik s to anomalijo. Na sl. 7.27 prikazuje rodovnik z retinoblastomom v primeru nepopolne penetracije.
Recesivne lastnosti se fenotipsko pojavijo le pri homozigotih za recesivne alele. Ti znaki so običajno

riž. 7.26. Rodovnik (X) z avtosomno dominantnim tipom dedovanja (brahidaktilija - B)

riž. 7.27.
riž. 7.28. Verjetnost, da bi imeli potomce recesivna lastnost iz različnih parov
najdemo pri potomcih fenotipsko normalnih staršev - nosilcev recesivnih alelov. Verjetnost pojava recesivnega potomstva je v tem primeru 25%. Če ima eden od staršev recesivno lastnost, bo verjetnost njegove manifestacije pri potomcih odvisna od genotipa drugega starša. Pri recesivnih starših bodo vsi potomci podedovali ustrezno recesivno lastnost (slika 7.28).
Za rodovnike z avtosomno recesivnim dednim vzorcem je značilno, da se lastnost ne pojavlja v vsaki generaciji. Najpogosteje se recesivni potomci pojavljajo pri starših z dominantno lastnostjo, verjetnost takšnega potomstva pa se pojavi v tesno sorodnih zakonih, kjer sta oba starša lahko nosilca istega recesivnega alela, pridobljenega od skupnega prednika. Primer avtosomno recesivnega dedovanja je rodovnik družine s psevdohipertrofična progresivna miopatija, v katerih so pogoste tesno povezane poroke (slika 7.29). Opozarja na širjenje bolezni v zadnji generaciji horizontalno.
Geni, ki se nahajajo na kromosomu X, ki jih nimajo

riž. 7.29. Rodovnik avtosomno recesivnega dedovanja (psevdohipertrofična progresivna miopatija) alelov v Y-kromosomu, predstavljenih v genotipih moških in žensk v različnih odmerkih. Ženska prejme svoja dva kromosoma X in ustrezne gene od očeta in matere, moški pa svoj edini kromosom X podeduje po materi. Razvoj ustrezne lastnosti pri moških določa en sam alel, ki je prisoten v njegovem genotipu, pri ženskah pa je posledica interakcije dveh alelnih genov. V zvezi s tem se v populaciji z različno verjetnostjo najdejo lastnosti, podedovane po X-vezanem tipu pri moških in ženskah.
S prevladujočim dedovanjem, vezanim na X, je ta lastnost pogostejša pri ženskah zaradi njihove večje sposobnosti sprejemanja ustreznega alela od očeta ali matere. Moški lahko to lastnost podedujejo le po materi. Ženske z dominantno lastnostjo jo prenašajo enako na svoje hčere in sinove, moški pa le na svoje hčere. Sinovi nikoli ne podedujejo prevladujoče lastnosti, povezane z X-om, od svojih očetov.
Primer te vrste dedovanja je rodovnik, opisan leta 1925 s folikularna keratoza - kožna bolezen spremlja izguba trepalnic, obrvi, las na glavi (slika 7.30). Značilen je težji potek bolezni pri hemizigotnih moških kot pri ženskah, ki so najpogosteje heterozigoti.
Pri nekaterih boleznih opazimo smrt moških hemizigotov v zgodnjih fazah ontogeneze. Potem naj bodo v rodovniku med obolelimi le ženske, pri katerih je razmerje med prizadetimi hčerami, zdravimi hčerkami in zdravimi sinovi 1:1:1. Moške dominantne hemizigote, ki ne umrejo v zelo zgodnjih fazah razvoja, najdemo pri spontanih splavih ali med mrtvorojenimi otroki. Za takšne značilnosti dedovanja pri ljudeh je značilna pigmentna dermatoza.
Značilna lastnost rodovnikov s to vrsto dedovanja je prevladujoča manifestacija lastnosti pri hemizigotnih moških, ki jo podedujejo po materah.

riž. 7.30. Rodovnik z dominantnim tipom dedovanja, vezanim na X (folikularna keratoza)

riž. 7.31. Rodovnik z X-vezanim recesivnim dedovanjem (hemofilija tipa A)
z dominantnim fenotipom, ki so nosilci recesivnega alela. Praviloma to lastnost podedujejo moški skozi generacijo od dedka po materi do vnuka. Pri ženskah se kaže le v homozigotnem stanju, za katerega se verjetnost poveča s tesno povezanimi porokami.
Najbolj znan primer recesivnega X-vezanega dedovanja je hemofilija. Dedovanje hemofilije tipa A je predstavljeno v rodovniku potomcev angleške kraljice Viktorije (slika 7.31).
Drug primer dedovanja te vrste je barvna slepota- določena oblika kršitve zaznavanja barv.
Prisotnost Y kromosoma samo pri moških pojasnjuje posebnosti Y-povezanega ali nizozemskega dedovanja lastnosti, ki je prisotna le pri moških in se prenaša po moški liniji iz roda v rod z očeta na sina.

riž. 7.32. Rodovnik z Y-vezanim (holandskim) tipom dedovanja
Ena lastnost, katere dedovanje, povezano z Y pri ljudeh, je še vedno predmet razprave hipertrihoza ušesa, ali prisotnost dlak na zunanjem robu ušesa. Domneva se, da poleg tega gena kratka roka Y kromosoma vsebuje gene, ki določajo moški spol. Leta 1955 so pri miši opisali transplantacijski antigen, vezan na Y, imenovan HY.
Morda je to eden od dejavnikov spolne diferenciacije moških spolnih žlez, katerih celice imajo receptorje, ki vežejo ta antigen. Antigen, vezan na receptor, aktivira razvoj spolne žleze moški tip(Glejte razdelke 3.6.5.2; 7.1.2).
Ta antigen je med evolucijo ostal skoraj nespremenjen in ga najdemo v telesu številnih živalskih vrst, vključno z
in oseba. Tako je dedovanje sposobnosti razvoja spolnih žlez po moškem tipu določeno s holandskim genom, ki se nahaja na Y kromosomu (slika 7.32).
Oseba kot predmet genetskega raziskovanja nima skoraj nobenih prednosti pred drugimi predmeti.
Nasprotno, obstajajo številne ovire, ki otežujejo preučevanje njegove genetike: 1) nemožnost poljubnega križanja v poskusu; 2) pozen začetek pubertete; 3) majhno število potomcev v vsaki družini; 4) nezmožnost izenačevanja življenjskih pogojev za potomce; 5) pomanjkanje natančne registracije manifestacije dednih lastnosti v družinah in odsotnost homozigotnih linij; 6) veliko število kromosomov; 7) in najpomembnejša težava pri preučevanju človeške genetike v kapitalistični družbi je družbena neenakost, ki otežuje uresničevanje človekovih dednih potencialov.
Kljub tem težavam je genetika razvila nekaj metod, ki omogočajo preučevanje dednosti in dedovanja pri ljudeh korak za korakom. Obstaja več raziskovalnih metod: genealoška, citogenetska, dvojčka, ontogenetska in populacijska.
Upoštevati je treba, da lahko katera koli lastnost, ne glede na to, ali gre za lastnost divjega tipa, torej nanaša na normo ali je povezana s katero koli boleznijo, služi kot model za preučevanje dednosti. Zaščita osebe pred dednimi boleznimi ali poškodbami njegove dednosti je enako pomembna kot ugotovitev dedovanja norme. Trenutno se genetske metode razvijajo predvsem v zvezi z morfološkimi lastnostmi, ki so genetsko dokaj jasno določene (brahidaktilija, albinizem, barvna slepota, pegavost kože in las itd.).
Genetsko preučevanje duševnih lastnosti še vedno ostaja problematično, saj zanje niso našli osnovnih kriterijev za lastnost v genetskem smislu. Skoraj vsi znaki človekove miselne in ustvarjalne dejavnosti so tako zapleteni in zapleteni ter močno pogojeni tudi z zunanjimi, vključno s socialnimi dejavniki, da je še vedno težko izvesti genetsko analizo teh lastnosti, čeprav je njihova dedna pogojenost nedvomna. .
Lahko rečemo, da je veliko večino lastnosti, ki so značilne za vrsto Homo sapiens, mogoče proučevati kot kvantitativne in kompleksne fiziološke lastnosti, torej lastnosti, ki v ontogenezi ne kažejo diskretnega značaja. Te lastnosti nadzoruje sistem genotipov (poligen). In dokler tega sistema ne razkrijemo, vsaj na primeru preprosto organiziranih organizmov, ostaja problem znakov vedenja nedostopen za genetsko analizo. Nasprotno, mutantne lastnosti, ki presegajo značilnosti lastnosti vrste, služijo kot dobri genetski modeli za preučevanje dednosti in dedovanja v normi.
Diskretnih mutantnih lastnosti ni mogoče obravnavati le kot patološke lastnosti, ki domnevno nimajo prilagoditvene vrednosti. Možno je, da je že sam videz osebe z razvitimi hemisferami možganske skorje, navpičnim položajem telesa in diskretno govorno signalizacijo posledica večjih mutacij. To močno podpira
kratko obdobje v človeški evoluciji, v katerem bi se majhne mutacije komaj nabrale v tolikšni količini in bi dale tako pomemben evolucijski učinek. Človek s smislom za naravo je "nenavadna" kot domača kokoš, ki namesto 10-15 znese 365 jajc na leto, ali krava rekorderka, ki namesto 600-700 kg da 16.000 kg mleka na leto.
Delitev lastnosti na normalne in mutantne v odnosu do ljudi in živali je nujna za poznavanje človekove evolucije in patoloških pojavov.
Celovitost vrstnih značilnosti ljudi in živali je določena s sistemom genotipov, ki se je razvil pod vplivom vseh selekcijskih dejavnikov v procesu evolucije. Zdi se, da so mutacije, ki so heterozigotne pri ljudeh, prav tako potrebne kot pri živalih, da jih ohranijo v populaciji.
Najnevarnejša stvar pri razvoju znanstvenih metod za preučevanje živali in človeka, predvsem njegovih sposobnosti, je antropomorfni moment, torej željno razmišljanje.
genealoška metoda
Analiza človeške dediščine na podlagi sestave rodovnika – rodoslovja je predlagal F. Galton.
genealoška metoda je študija dedovanja človeških lastnosti glede na rodovnike (rodovnik). Ta metoda velja, če so znani neposredni sorodniki - predniki lastnika dedne lastnosti (probanda) po materini in očetovi liniji v več generacijah in je v vsaki generaciji zadostno število potomcev ali v primeru, ko so podatki o zadostnem številu različnih družin, da se razkrije podobnost rodovnikov. Podatki o nizu podobnih rodovnikov so podvrženi statistični obdelavi.
Najbolj razširjen sistem za označevanje človeških rodovnikov je leta 1931 predlagal G. Yust.
Na podlagi velikega števila analiziranih družin se sestavijo rodovniki in naredijo matematični izračuni glede na vrsto dedovanja določene lastnosti – dominantno ali recesivno, pogosto in redko pojavljajoče se mutacije, spolno vezane ali ne, itd. ne bomo se dotaknili uporabe matematične metode pri tej analizi, omenimo le, da celotna formalna analiza temelji na elementarnih genetskih vzorcih dedovanja.
Rodovniški vzorci dedovanja prevladujočega avtosomnega gena, ki določa lastnost, kot je bolezen (hondrodistrofični pritlikavost, bulozna epidermoliza – sposobnost kože, da ob manjših poškodbah tvori velike mehurje, retinoblastom ipd.), ali morfološka napaka, kot npr. kratki prsti (brahidaktilija - odsotnost dveh distalnih falang v prstih).

Dedovanje lastnosti, ki jih določajo recesivni geni (recesivno dedovanje), je pri sestavljanju rodovniških kart analizirano nekoliko bolj zapleteno.
Na primer, dva v družini je pojav dveh bolnih otrok enak zmnožku verjetnosti, to je 0,25 X 0,25, torej 6,25%.
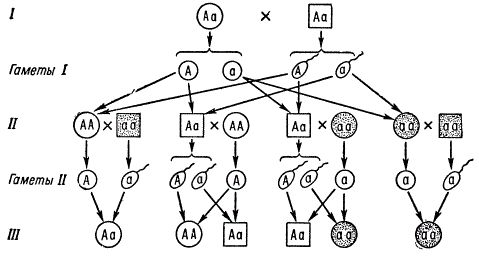
Pogosto pojavljajoči se recesivni avtosomni geni, pod pogojem, da se njihovi nosilci (aa) lahko poročijo in rodijo potomce, bodo v visoki koncentraciji v populaciji. V tem primeru postanejo zakonske zveze aa X Aa zelo verjetne, pri potomcih, od katerih bo dedovanje te lastnosti posnemalo dedovanje po prevladujočem tipu 1: 1. Vendar je ob poznavanju vrste dedovanja in manifestacije teh in drugih genov, tudi v primeru majhnih družin, vendar z zadostnim številom takšnih družin, mogoče ugotoviti pravo naravo dedovanja.
Dedovanje genov, ki so v celoti vezani na spol, torej locirani v nehomolognih segmentih, in delno vezani na spol - lokalizirani v homolognih segmentih X- in Y-kromosomov, sledi vzorcem, uveljavljenim za spolne kromosome. Za dominantno in recesivni geni to dedovanje bo različno opredeljeno glede na to, kje se gen nahaja – v homolognem ali nehomolognem segmentu X- in Y-kromosoma in kako se prenaša. Torej je prevladujoči gen, ki povzroča prepletene prste, ki se nahaja v nehomolognem segmentu Y kromosoma, podedovan od očetov in se kaže le pri moških.
Za delno spolno povezane dominantne gene, ki se nahajajo v homolognih segmentih spolnih kromosomov, je analiza nekoliko težja, a tudi možna. Primer spolno povezanega dedovanja recesivne lastnosti je dedovanje hemofilije. Obstaja diskontinuiteta v prenosu te lastnosti v generacijah; prizadeti moški so potomci zdravih mater, ki so bile heterozigotne za gen; ženske s hemofilijo so lahko potomke bolnega očeta in bolne ali zdrave matere.

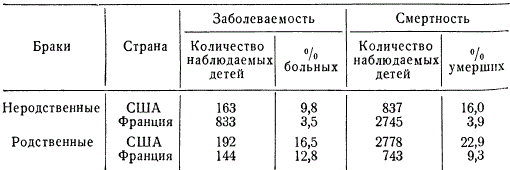
Pri ljudeh so odkrili približno 50 spolno povezanih recesivnih genov. Zanimivo je, da jih približno polovica povzroča očesne bolezni. Že dolgo je znano, da je stopnja prenosa dednih lastnosti v sorodnih (inbreeding) in nepovezanih zakonih (outbreeding) različna. Potem ko je genetika vzpostavila vzorce več pogosta manifestacija recesivnih genov pri parjenju v sorodstvu, ni treba obširno dokazovati škode sorodstvenih zakonov. Višji kot je koeficient inbreedinga, večja je verjetnost pojava dednih bolezni v generacijah. IN različne države med različni narodi in družbenih razredov, pa tudi v različnih obdobjih sorodne poroke (med bratranci, bratranci in sestrami) se pojavljajo z različno pogostostjo. Tako na primer v vaseh na otokih Fidži število sorodnih porok doseže 29,7%, v nekaterih kastah Indije - 12,9, na Japonskem (Nagasaki) - 5,03, na Nizozemskem - 0,13-0,159, na Portugalskem - 1 40, v ZDA (Baltimore) - 0,05 % itd. Odstotek sorodstvenih porok se v različnih regijah iste države razlikuje, odvisno od načina življenja.
Škodljivost sorodstvenih porok je v posameznih rodovnikih komaj opazna, s primerjalno statistično analizo bolezni in umrljivosti pa se kaže povsem očitno.
Osupljiv primer odkrivanja recesivnega gena v sorodstveni zakonski zvezi.
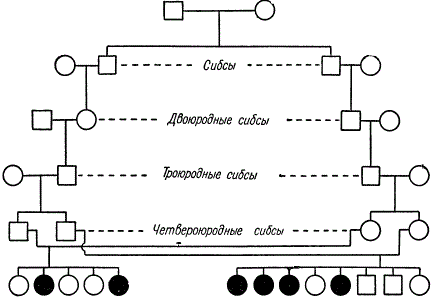
V tem rodovniku se sorodstvo ohranja s poroko bratov in sester (bratov in sester) različnih stopenj sorodstva. Iz dveh sorodstvenih zakonov (četrti bratranci in sestre) so se v eni družini pojavili 4 od 8 otrok, v drugi pa 2 od 5 otrok, ki trpijo za dednim amavrotičnim idiotizmom. K. Stern predlaga, da je eden od dveh skupnih prednikov teh linij ta recesivni gen posredoval skozi tri generacije na vsakega od štirih staršev.
Včasih bolezen in umrljivost otrok iz sorodnih zakonov presegata za 20-30 % tiste iz nepovezanih zakonov. Očitno je, da je vzrok obravnavanega pojava genetski, in sicer: velika verjetnost manifestacije dedne bolezni in umrljivost kot posledica homozigotizacije recesivnih genov, ki določajo fiziološke insuficience in umrljivost (smrtonosni in polletalni geni).
Rodoslovna metoda je torej zelo dragocena metoda, vendar je njen pomen pri raziskovanju toliko večji, kolikor natančnejši in globlje so sestavljeni rodovniki. Z rastjo civilizacije in natančnejšo registracijo rodovnikov se bo vloga te metode v človeški genetiki povečala.
dvojna metoda
dvojčka imenovani potomci, sestavljeni iz istočasno rojenih posameznikov v samskih živalih (ljudje, konji, govedo, ovce itd.).
Dvojčka sta lahko enojajčna ali bratska.
identični oz enojajčna dvojčka(OB) se razvijejo iz enega jajčeca, oplojenega z eno semenčico, ko dva ali več zarodkov nastaneta iz zigote namesto enega zarodka (poliembrion). Ker mitotična delitev zigote daje dva enako dedna blastomera, morajo biti enojajčni dvojčki, ne glede na to, koliko se jih razvije, dedno identični in istega spola. Ta pojav je primer aseksualnega ali bolje rečeno vegetativnega razmnoževanja živali.
bratska dvojčka(RB) se razvijejo iz istočasno ovuliranih različnih jajčec, oplojenih z različnimi spermatozoidi. In ker lahko nosijo različna jajčeca in sperme različne kombinacije genov, potem so lahko dvojčki dedno tako različni kot otroci istega zakonskega para, rojenega leta drugačen čas. Bratovski dvojčki so lahko istega (RBR) ali različnega spola (RBR).
Pogosteje se v literaturi namesto izraza "bratski dvojčki" (RB) uporablja izraz "bratski dvojčki" (DB), saj so dvojčki pogostejši. Vendar pa izraz "bratovski dvojčki" bolje poudari razliko med OB in RB; enojajčni dvojčki so tudi bolj verjetno rojeni kot dvojčki.
Sodeč po podatkih, pridobljenih o sesalcih, lahko obstaja več hipotez, ki pojasnjujejo nastanek OB pri ljudeh:
- razhajanje blastomerov med prvo delitvijo zigote in ločen razvoj zarodka od teh blastomerov;
- ločitev skupine celic v fazi blastociste (pred gastrulacijo);
- ločitev zarodkov v zgodnji fazi gastrulacije. Najbolj verjeten način je drugi.

Število dvojčkov pri enem rojstvu pri človeku je različno: najpogosteje so dvojčki, redkeje so trojčki, še redkeje so četverčki, zelo redko pa je pet. Po besedah I. I. Kanaeva so v zadnjih 150 letih v Združenih državah Amerike našli štiri primere peterčkov in dva primera v Kanadi. Dejstvo rojstva OB - petih deklet, ki so preživele do odraslosti - je znano v družini kanadskega kmeta Dionne (1934). Izračunano je, da se petero rodi enkrat na 54.700.816 rojstev, zobniki - pri 4.712 milijonih rojstev so sedmice znane le kot izjema. V povprečju je rodnost dvojčkov 1% z nihanji v območju 0,5-1,5%. Dvojčki so manj sposobni preživeti, zato je njihovo število ob rojstvu manjše kot ob spočetju, v odrasli dobi pa manjše kot ob rojstvu.
Izračun pogostosti OB glede na RB temelji na teoretičnem razmerju istospolnih in nasprotnospolnih parov RB ob rojstvu dvojčkov: 25%♀♀ + 50%♀♂ + 25%♂♂ ženska) bo dalo razliko v številu OB parov, ki se v povprečju giblje od 21 do 33,4 % vseh dvojčkov.
Za uporabo dvojčkov v genetskih študijah je zelo pomembno natančno diagnosticirati vrsto AB in vrsto RP. Diagnoza se postavi na podlagi naslednjih meril:
- OB morajo biti istega spola, RB so lahko istega ali različnih spolov;
- OB imajo praviloma en skupni horion, RB - različni korioni;
- recipročna presaditev tkiva v OB je tako uspešna kot avtotransplantacija, v RB je nemogoča;
- prisotnost podobnosti (konkordanca) v OB in neskladnost (diskordanca) v RB v mnogih pogledih.
Za diagnozo je treba izbrati znake, ki so očitno podedovani in so najmanj dovzetni za spremembe pod vplivom okoljskih dejavnikov; taki znaki vključujejo krvne skupine, pigmentacijo oči, kože in las, reliefnost kože (prstni odtisi, dlani, stopala itd.). Če ena ali dve od teh lastnosti razkrivata razliko med dvojčki, so praviloma RB.
Vse dvomljive primere diagnoze dvojčkov lahko povzroči bodisi motnja v razvoju enega od OB partnerjev bodisi podobnost staršev na več načinov. Vendar pa je slednje izjemno redko. Opozoriti je treba, da razvojno motnjo enega od OB partnerjev običajno pojasnimo z neenakomernim delovanjem dejavnikov. intrauterino življenje in pojav somatskih mutacij v zgodnjih fazah embrionalnega razvoja, pred nastankom organov. Različne vrste genskih in kromosomskih preureditev, monosomija in druge mutacije, ki se pojavijo pri enem od partnerjev, lahko povzročijo pomembne razlike v fenotipu OB. Zato je treba v zgodnji embriogenezi upoštevati možnost somatskih mutacij v OB.
Glede na posplošitve I. I. Kanaeva, ki jih je postavila v njegovi odlični monografiji, bistvo dvojna metoda v genetiki se zmanjša na naslednje določbe:
1) par OB ima identično kombinacijo, par RB ima različne kombinacije genotipov staršev;
2) za oba partnerja enega para OB se lahko zunanje okolje izkaže za enako, za drugega pa različno. Če imajo OB partnerji izkušnje drugačen vpliv, potem bo to vodilo do razlike v paru. Zato so lahko pari z identičnimi okolji znotraj para in različnim okoljem znotraj para.
Primerjava OB z enakim okoljem z OB z drugačnim okoljem odpira možnost presoje vloge vpliva okolja na razlike znotraj parov pri dvojčkih skozi vse življenje. Primerjava OB z istim okoljem in RB z istim okoljem vam omogoča, da ugotovite vlogo dednega faktorja. Tovrstna študija je izvedena na velikem vzorcu in statistično obdelana.
Na podlagi razlike v genetskem izvoru OB in RB sledi, da če ni razlik v OB za iste lastnosti in obstajajo takšne razlike v RB, potem je očitno, da so te razlike v lastnostih pri slednjih posledica dedni dejavniki. Če pa se pri eni in drugi vrsti dvojčkov pojavijo razlike v istih lastnostih znotraj para, potem je očitno, da jih lahko povzročijo okoljski dejavniki. Iz podatkov o neskladju v OB in RB za številne morfološke značilnosti je razvidno, da se znotrajparska razlika v RB pojavlja večkrat pogosteje kot pri OB.

Predstavljeni so nekateri podatki S. Reeda o primerjalni pogostnosti patologije pri drugem partnerju v primeru bolezni enega od dvojčkov.

Odstotek prikazuje pogostost skladnosti bolezni pri dveh vrstah dvojčkov, kaže, da če je en partner zbolel za eno od indiciranih bolezni, je verjetnost za drugo bolezen pri OB veliko večja kot pri RB. VP Efroimson, ki analizira podatke o pogostosti kontorcijskih parov, povsem pravilno poudarja, da se visoka dedna nagnjenost OB k boleznim kaže v prisotnosti provocirajočega dejavnika; brez tega bo ta odstotek precej nižji.
Metoda dvojčka omogoča z največjo natančnostjo določitev dedne nagnjenosti osebe k številnim boleznim in lastnostim. Z drugimi metodami je zelo težko ali skoraj nemogoče preučiti številne nalezljive in tumorske bolezni, vnetja kože in različnih organov ter značilnosti normalnega človekovega živčnega delovanja.
Pri uporabi metode dvojčkov je treba upoštevati pogoje skupne in ločene vzgoje v življenju partnerjev, socialne razmere, v katerih se nahajajo, itd. preučevane gene in izolirati vlogo okolja pri določanju variabilnosti. preučenih lastnosti.
Citogenetska metoda
Citogenetska metoda v človeški genetiki se običajno imenuje citološka analiza kariotipa osebe v normalnih in patoloških stanjih.
Bolj pravilno je to metodo imenovati citološka in ne citogenetska, saj je genetska analiza s križanjem pri ljudeh izključena, nosilci kromosomskih motenj, če preživijo, pa so običajno neplodni. Občasno pa je pri nekaterih kromosomskih motnjah možno kombinirati citološko metodo z genealoško in ugotoviti povezavo med fenotipskim učinkom in določeno vrsto kromosomskih sprememb. Zaradi teh okoliščin je mogoče obdržati izraz »citogenetska metoda«, ki je v literaturi sprejet pri preučevanju človeške genetike. V primerih, ko se takšen paralelizem ne preučuje, uporaba tega izraza ni dovoljena.
Citogenetska metoda se uporablja za raziskovanje različnih vrst heteroploidije in kromosomskih preureditev v človeških somatskih tkivih, ki povzročajo različna fenotipska odstopanja od norme.
Najpogosteje se ta metoda uporablja v tkivni kulturi. Omogoča upoštevanje velikih kromosomskih anomalij, ki se pojavljajo tako v zarodnih kot v somatskih celicah. Izkazalo se je, da se pri ljudeh, pa tudi pri živalih, zaradi neločitve avtosomov in spolnih kromosomov med mejozo pogosto pojavijo trisomije in monosomije za različne pare kromosomov. Trisomijo in monosomijo za spolne kromosome pri ljudeh odkrijemo na podlagi analize spolnega kromatina.
V sorazmerno dolgem individualnem razvoju človeka se v celicah različnih tkiv kopičijo kromosomske anomalije (kromosomske preureditve, pa tudi sprememba števila kromosomov). Telesna tkiva so raznolike populacije genetsko različnih celic, v katerih se s starostjo povečuje koncentracija celic s patološkimi jedri. V tem primeru citogenetska metoda omogoča preučevanje staranja tkiva na podlagi preučevanja celičnih struktur v starostni dinamiki "populacije" somatskih in generativnih tkiv.
Ker je pogostost pojavljanja kromosomskih nepravilnosti odvisna od vpliva različnih mutagenov na telo (ionizacija, kemična sredstva - farmakološki pripravki, plinska sestava okolja itd.), citogenetska metoda omogoča ugotavljanje mutagenega učinka okoljski dejavniki na osebo.
Uporaba citogenetske metode se je še posebej razširila v povezavi z odkrivanjem vzrokov številnih telesnih in duševnih bolezni – tako imenovanih kromosomskih bolezni.
Obstaja več človeških bolezni, kot so Klinefelterjeva bolezen, Shereshevsky-Turnerjeva bolezen, Downova bolezen in druge, katerih vzroki so dolgo ostali neznani, dokler pri takih bolnikih s citološkimi metodami niso odkrili kromosomskih nepravilnosti.
Za bolne moške s Klinefelterjevim sindromom je značilna nerazvitost spolnih žlez, degeneracija semenskih tubulov, duševna zaostalost, nesorazmerna rast okončin itd. Shereshevsky-Turnerjev sindrom se pojavlja pri ženskah. Kaže se v zapozneli puberteti, nerazvitosti spolnih žlez, odsotnosti menstruacije, neplodnosti, nizki rasti in drugih patoloških znakih.
Izkazalo se je, da sta oba sindroma pri potomcih posledica nedisjunkcije spolnih kromosomov med nastajanjem starševskih gamet. Zaradi neločevanja kromosomov X pri ženskem homogametnem) spolu se lahko med mejozo pojavijo gamete z dvema X kromosomoma, to je XX + 22 avtosomi, in brez kromosomov X, to je 0 + 22; pri moškem (heterogametskem) spolu sta gameti XY + 22 oziroma 0 + 22. V primeru oploditve takšnih jajčec z normalnimi semenčici (X + 22 ali Y + 22) se lahko tvorijo naslednji razredi zigot: XXX + 44, 0X + 44 in XXY + 44, 0Y+44.
Iz tega sledi, da je število kromosomov v zigotah drugačnega izvora lahko variira od 47 do 45, posamezniki 0Y + 44 pa očitno ne preživijo, saj jih nikoli niso našli. Kromosomski niz XXY + 44 je neločljiv pri moškem s Klinefelterjevim sindromom (moška interseksualnost), kromosomski niz X0 + 44 in XXX + 44 najdemo pri ženskah s Shereshevsky-Turnerjevim sindromom.
Pri nadaljnji analizi bolnikov z različnimi sindromi se je izkazalo, da lahko zaradi nerazhajanja spolnih kromosomov pride do različnih vrst kromosomskih anomalij, zlasti polisomij. Obstajajo na primer moški s takšnimi nizi kromosomov: XX Y, XXX Y, XXXX Y in ženske - XXX, XXXX.
Posebnost vloge spolnih kromosomov pri določanju spola pri ljudeh v primeru njihove neločitve, v nasprotju z Drosophilo, se je pokazala v tem, da kromosomski niz XX Y vedno določa moški spol, niz X0 pa določa ženski spol. Hkrati pa povečanje števila kromosomov X v kombinaciji z enim Y kromosomom ne spremeni definicije moškega, ampak samo okrepi Klinefelterjev sindrom. Trisomija ali polisomija na kromosomu X pri ženskah pogosto povzroča tudi bolezni, podobne Shereshevsky-Turnerjevemu sindromu.
Bolezni, ki jih povzroča kršitev normalnega števila spolnih kromosomov, se diagnosticirajo s citološko metodo - analizo spolnega kromatina. V primerih, ko imajo moška tkiva normalen nabor spolnih kromosomov - XY, spolnega kromatina v celicah ni. Pri normalnih ženskah - XX - ga najdemo v obliki enega samega telesa. Pri polisomiji kromosoma X pri ženskah in moških je število teles spolnega kromatina vedno za eno manjše od števila kromosomov X, tj. nx \u003d n X - 1. Tako je v celicah moških s Klinefelterjevim sindromom, ko je XX Y rekrutirano, obstaja en telesni spolni kromatin, z nizom XXXY - dva, z nizom XXXXY - tri; pri ženskah s Shereshevsky-Turnerjevim sindromom: X0 - brez telesa, XXX - dve telesi, XXXX - tri telesa s spolnim kromatinom itd. Predpostavlja se, da je v vsaki takšni zigoti genetsko aktiven samo en od kromosomov X. Preostali kromosomi preidejo v heteropiknotično stanje v obliki spolnega kromatina.
Razlogi za ta vzorec niso pojasnjeni, vendar se domneva, da je povezan z izravnavo delovanja genov spolnih kromosomov pri hetero- in homogametnem spolu.
Kot vemo, se lahko neločitev kromosomov pojavi ne le v mejozi, temveč tudi v somatskih celicah med celotno embriogenezo živali, začenši od prvih cepitev jajčeca. Zaradi slednjega se lahko med ljudmi s kršitvijo razhajanja spolnih kromosomov pojavijo bolni ženski mozaiki in moški mozaiki. Opisani so na primer mozaiki naslednjih vrst: dvojni: X0/XX, X0/XXX in X0/XY, X0/XYY, trojni: X0/XX/XXX, XX/X0/XY, pa tudi štirikratni mozaiki, kadar somatske celice enega človeka vsebujejo štiri različne sklope kromosomov.
Poleg obravnavane vrste bolezni, ki jo povzroča sprememba števila spolnih kromosomov v zigoti, so lahko kromosomske bolezni posledica neločitve avtosomov in različnih vrst kromosomskih preureditev (translokacije, delecije). Tako na primer pri otrocih s prirojeno idiotnostjo - Downovo bolezen, ki jo spremlja nizka rast, široka okrogel obraz, tesno razporejena z ozkimi palpebralnimi razpokami in napol odprtimi usti, je bila odkrita trisomija na 21. kromosomu. Ugotovljeno je bilo, da je pojavnost Downove bolezni pri novorojenčkih odvisna od starosti mater.
S prirojenimi kromosomskimi nepravilnostmi so povezane številne bolezni. Zato je citogenetska metoda velikega pomena v etiologiji človeških bolezni.
populacijska metoda
populacijska metoda vam omogoča preučevanje porazdelitve posameznih genov ali kromosomskih nepravilnosti v človeški populaciji.
Metoda populacije temelji na matematičnih metodah. Za analizo genetske strukture populacije je treba pregledati velik vzorec, ki mora biti reprezentativen – objektivno odražati celotno generalno populacijo, torej celotno populacijo kot celoto. V pregledanem vzorcu je razporeditev oseb ugotovljena po ustreznih jasno opredeljenih fenotipskih razredih, med katerimi so razlike dedno določene. Nato se na podlagi najdenih fenotipskih frekvenc določijo frekvence genov.
Na podlagi poznavanja frekvenc genov je mogoče analizirano populacijo opisati v skladu s formulo Hardy-Weinberg in vnaprej predvideti verjetno naravo cepitve pri potomcih posameznikov, ki pripadajo enemu ali drugemu fenotipskemu razredu. Preučevanje genskih frekvenc je pomembno za ocenjevanje posledic sorodstvenih zakonov, pa tudi za razjasnitev genetske zgodovine človeške populacije kot celote.
Pogostost porazdelitve v populacijah različnih anomalij se izkaže za različno; medtem ko je velika večina ustreznih recesivnih alelov predstavljenih v heterozigotnem stanju.
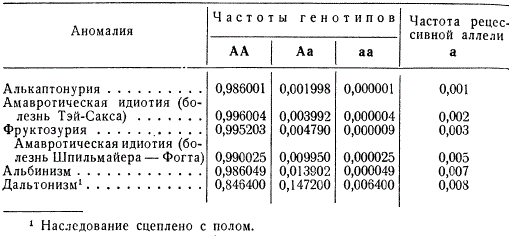
Tako je približno vsak stoti prebivalec Evrope heterozigoten za gen amavrotskega idiotizma (Spielmeier-Vogtova bolezen), medtem ko le 25 ljudi od 1 milijona homozigotnih zboli za to bolezen v adolescenci. Albini v evropskih državah se pojavljajo s pogostnostjo 1 na 20.000, čeprav je heterozigotno stanje tega alela lastno vsakemu sedemdesetemu prebivalcu.
Nekoliko drugačna situacija je v primeru anomalij, ki so podedovane spolno povezane, primer za to je barvna slepota - barvna slepota, ki jo očitno nadzoruje število alelov, razporejenih na dva tesno povezana lokusa na X kromosomu. Med moško populacijo pogostost barvne slepote (q) ustreza skupni pogostosti recesivnih alelov in je bila na primer v Moskvi v 30-ih letih, po podatkih RI Serebrovskaya, 7 %, medtem ko je med ženskami populacije iste populacije je bila barvna slepota le 0,5 % (q 2), vendar v heterozigotnem stanju približno 13 % žensk nosi alele, ki povzročajo barvno slepoto.
Kot smo povedali zgoraj, je glede na genealoško metodo verjetnost pojava recesivnih homozigotov pri potomcih lahko različna, če se poročijo osebe z različnimi stopnjami sorodstva. Torej, za zakonce, ki sta drug z drugim v razmerju bratranci in sestre, verjetnost, da imamo otroke homozigotne za recesivni alel, pogost v populaciji s frekvenco q, ne bo več q 2, ampak velika vrednost, in sicer q / 16 (1 + 15q).
To je posledica dejstva, da če je eden od skupnih prednikov takšnih zakoncev - babica ali dedek - nosil recesivni alel v heterozigotu, se bo ta alel z verjetnostjo 1/16 prenesel na oba bratranca.
Škodljivi učinki sorodstvenih porok so še posebej očitni pri izoliranih populacijah omejene velikosti, t.i. izolati. Izolat je skupina posameznikov v populaciji, ki se večinoma poročijo s posamezniki svoje skupine in so zato označeni s pomembnim koeficientom sorodstva. Takšni izolati so lahko ločene izolirane vasi, skupnosti itd. Znotraj izolata so bolj verjetne sorodne poroke (inbreeding), zakonca pa imajo večjo verjetnost, da nosijo iste mutantne gene, kar ima za posledico povečano verjetnost manifestacije recesivnih alelov. v homozigotnem stanju. Različni izolati imajo različne koncentracije podobnih ali različnih genov.
Na Marianskih otokih in otoku Guam je umrljivost lokalnega prebivalstva zaradi amiotrofične lateralne skleroze (povezane s poškodbami celic sprednjih rogov hrbtenjače) več kot 100-krat večja od umrljivosti zaradi te bolezni v drugih državah. držav. V južni Panami, v provinci San Blas, je zelo pomemben del plemena Carib Kuna albino, ki se tukaj pojavlja v vsaki generaciji. V eni vasi ob reki. Rhone v Švici je med 2200 prebivalci več kot 50 gluhonemih, 200 pa jih ima nekaj okvar sluha. Po vsej verjetnosti ima v vseh takih primerih močnega povečanja koncentracije posameznih alelov določeno vlogo genetski drift, neenakomerno razmnoževanje v preteklosti posameznih družin, rodov in zmanjšanje migracij.
Z rastjo civilizacije in razvojem proizvodnih sil družbe se število izolatov zmanjšuje, njihov pomen za celotno prebivalstvo pa se zmanjšuje. Vendar še vedno obstajajo.
Poznavanje genskih frekvenc, kot je bilo že omenjeno, omogoča napovedovanje narave cepitve pri potomcih posameznih fenotipskih razredov starševskih posameznikov.
Na podlagi formule Hardy-Weinberg je mogoče dokazati, da je treba pri monogenem dedovanju delitev po fenotipu pri potomcih dominantnih mater izvesti v razmerju p (1 + pq) dominantnih proti p recesivnim ali (l + pq). ): q 2; pri potomcih recesivnih mater mora biti fenotipska cepitev pq 2: q 3 ali p: q.
Vzemimo primer. V eni študiji je bila pri preučevanju faktorja Rh frekvenca recesivnega alela rh v populaciji 0,4, pogostnost dominantnega alela Rh pa 0,6. Zato je bilo pričakovati, da bo pri potomcih Rh-pozitivnih mater pogostnost Rh-pozitivnih otrok (Rh +) približno 7,8-krat večja od pogostnosti Rh-negativnih otrok (Rh -); pri potomcih Rh-negativnih mater bo ustrezen presežek 1,5-krat.
Dejanska razmerja v anketiranem vzorcu so bila:
- v prvem primeru 1475 Rh + : 182 Rh - ali 8,1: 1,
- v drugem primeru 204 Rh + : 129 Rh - ali 1,6: 1.
Tako se opaženi rezultati delitve zelo dobro ujemajo s teoretično pričakovanimi rezultati, predvidenimi iz analize genske frekvence.
Populacijska analiza polimorfizma po krvnih skupinah je zanimiva, ker pomaga razumeti dinamiko genetske strukture različnih populacij in pomaga pri prepoznavanju odnosov med njimi.
Različne populacije se bistveno razlikujejo po svoji genetski strukturi, zlasti po krvnih skupinah. Hkrati je mogoče zaslediti nekaj precej jasnih vzorcev. Če je koncentracija alela I B najvišja v regiji Indije in Kitajske, potem vzhodno in zahodno od te regije pri avtohtonih prebivalcih Amerike in Avstralije postopoma pada na nič. Hkrati pri ameriških Indijancih (ter staroselcih Avstralije in Polinezije) koncentracija alela I 0 doseže maksimum. Alel I A je redek pri avtohtonem prebivalstvu Amerike, pa tudi v Indiji, Arabiji, tropski Afriki in Zahodni Evropi.
Za razlago teh razlik v genetski strukturi populacij je bila pred kratkim predlagana hipoteza, po kateri sta odločilni dejavnik pri selekciji glede na krvne skupine sistema AB0 epidemije kuge in črnih koz. Povzročitelj kuge Pasteuvella pestis, ki ima lastnost antigena 0, je najbolj škodljiv za ljudi s krvno skupino 0, saj takšni posamezniki v primeru okužbe ne morejo proizvesti dovolj protiteles. Iz podobnega razloga je virus črnih koz najbolj nevaren za ljudi s krvno skupino A. Tam, kjer je divjala kuga (Indija, Mongolija, Kitajska, Egipt), je prišlo do intenzivne eliminacije alela I 0 in kjer so bile črne koze še posebej razširjene ( Amerika, Indija, Arabija, tropska Afrika) je bil najprej izločen alel 1 A. Na območjih Azije, kjer sta bili endemična kuga in črne koze, je bil alel 1 c deležen največje frekvence.
V 5. poglavju smo pregledali monogeno dedovanje anemije srpastih celic zaradi cepitve alelov S. rezultat je sistem uravnoteženega dednega polimorfizma.
Tako v obeh primerih analize polimorfizma po krvnih skupinah in srpastocelične anemije vidimo, kako nam uporaba populacijske metode omogoča razkrivanje genetske strukture človeških populacij.
ontogenetska metoda
ontogenetska metoda vam omogoča, da ugotovite prenos recesivnih alelov v heterozigotnem stanju in kromosomske preureditve po fenotipu.
Genetska osnova za manifestacijo recesivnih genov v heterozigotnem stanju je očitno nepopoln blok v verigi sinteze enega ali drugega presnovka, ki ga povzroča delovanje prevladujočega alela tega gena.
Znano je, da se nekatere dedne bolezni ne kažejo le pri osebah, homozigotnih za alele, ki povzročajo bolezen, temveč v izbrisani obliki tudi pri heterozigotih. Zato se trenutno intenzivno razvijajo metode za določanje heterozigotnega nosilca v ontogenezi. Torej, heterozigoten nosilec fenilketonurije (povečana vsebnost fenilalanina v krvi se določi z dodatnim dajanjem fenilalanina in naknadnim določanjem njegove (ali tirozina) ravni v krvni plazmi. Prisotnost heterozigotnosti za ta alel ugotavljamo z povišana vsebina fenilalanin. Običajno (tj. pri homozigotih za dominantni alel) se raven fenilalanina ne spremeni. Običajno kri vsebuje encim katalazo, ki je nujen za presnovo ogljikovih hidratov, vendar obstaja gen, ki v homozigotnem stanju povzroči odsotnost katalaze. Homozigotni nosilci tega gena imajo bolezen akatalazemijo - motnjo presnove ogljikovih hidratov. Heterozigoti so vmesni v aktivnosti katalaze brez velikega prekrivanja med dominantnimi in recesivnimi homozigoti.
Aktivnost katalaze lahko natančno identificira heterozigotne in homozigotne nosilce alela atalaze med bližnjimi sorodniki in starši.
Heterozigotno prenašanje alela, ki določa Duchennovo mišično distrofijo, testiramo z aktivnostjo kreatin fosfokinaze. Zdaj so bili razviti podobni testi za 40 dednih bolezni, določenih z recesivnimi aleli.
Trenutno so ontogenetsko metodo obogatile biokemične, imunološke in molekularne raziskovalne metode, ki so opisane v številnih posebnih priročnikih.
Pomen ontogenetske metode je očiten za ugotavljanje nosilnega stanja recesivnega gena v heterozigotnem stanju pri sorodnikih družine, v kateri se pojavlja dedno bolan otrok. Diagnostika v ontogenezi je pomembna za izračun verjetnosti pojava dedno bolnih potomcev v sorodnih in mešanih zakonih. Ker postane testiranje heterozigotnih nosilcev lažje, bo treba to metodo uvesti za svetovanje parom o možnosti bolezni pri njihovih otrocih, pa tudi za preučevanje širjenja mutacij v populacijah.
Če najdete napako, označite del besedila in kliknite Ctrl+Enter.
Osnova te metode je sestavljanje in analiza rodovnikov. Ta metoda se od antičnih časov do danes pogosto uporablja v konjereji, selekciji dragocenih linij goveda in prašičev, pri pridobivanju čistokrvnih psov, pa tudi pri vzreji novih pasem kožuharjev. Človeška rodoslovja so bila sestavljena skozi več stoletij v zvezi s vladajočimi družinami v Evropi in Aziji.
Kot metodo preučevanja človeške genetike se je genealoška metoda začela uporabljati šele v začetku 20. stoletja, ko je postalo jasno, da lahko analiza rodovnikov, v katerih se zasleduje lastnost (bolezen) iz roda v rod, nadomesti hibridološko metoda, ki je za ljudi dejansko neuporabna.
Pri sestavljanju rodovnikov je vir oseba - proband, katerega rodovnik se preučuje. Običajno je to bodisi bolnik ali nosilec določene lastnosti, katere dedovanje je treba preučiti. Pri sestavljanju rodoslovnih tabel se uporabljajo simboli, ki jih je leta 1931 predlagal G. Yust (slika 6.24). Generacije so označene z rimskimi številkami, posamezniki v dani generaciji so označeni z arabskimi številkami.
riž. 6.24. Simboli pri pripravi rodovnikov (po G. Yustu)
S pomočjo genealoške metode je mogoče ugotoviti dedno pogojenost proučevane lastnosti, pa tudi vrsto njenega dedovanja (avtosomno dominantno, avtosomno recesivno, X-vezano dominantno ali recesivno, Y-povezano). Pri analizi rodovnikov za več lastnosti je mogoče razkriti povezano naravo njihovega dedovanja, ki se uporablja pri sestavljanju kromosomskih kart. Ta metoda omogoča preučevanje intenzivnosti procesa mutacije, oceno ekspresivnosti in penetracije alela. Široko se uporablja v medicinskem genetskem svetovanju za napovedovanje potomcev. Vendar je treba opozoriti, da postane genealoška analiza veliko bolj zapletena, ko imajo družine malo otrok.
Rodovniki pri avtosomno dominantnem dedovanju. Za avtosomni tip dedovanja je na splošno značilna enaka verjetnost pojava te lastnosti pri moških in ženskah. To je posledica enakega dvojnega odmerka genov, ki se nahajajo v avtosomih vseh predstavnikov vrste in so jih prejeli od obeh staršev, in odvisnosti lastnosti v razvoju od narave interakcije alelnih genov.
Kadar lastnost prevladuje pri potomcih starševskega para, kjer je vsaj eden od staršev njen nosilec, se kaže z večjo ali manjšo verjetnostjo, odvisno od genetske konstitucije staršev (slika 6.25).

riž. 6.25. Verjetnost pojava potomcev s prevladujočo lastnostjo iz različnih zakonskih parov (/- III)
Če analiziramo lastnost, ki ne vpliva na sposobnost preživetja organizma, so lahko nosilci prevladujoče lastnosti tako homo- kot heterozigoti. V primeru prevladujočega dedovanja neke patološke lastnosti (bolezni) homozigoti praviloma niso sposobni preživeti, nosilci te lastnosti pa so heterozigoti.
Tako se pri avtosomno dominantnem dedovanju lahko lastnost enako pojavlja pri moških in ženskah in jo je mogoče zaslediti z zadostnim številom potomcev v vsaki generaciji vzdolž vertikale. Pri analizi rodovnikov se je treba spomniti možnosti nepopolnega prodora prevladujočega alela zaradi interakcije genov ali okoljskih dejavnikov. Indeks penetracije lahko izračunamo kot razmerje med dejanskim številom nosilcev lastnosti in številom pričakovanih nosilcev te lastnosti v dani družini. Ne smemo pozabiti tudi, da se nekatere bolezni ne pojavijo takoj po rojstvu otroka. Številne bolezni, podedovane po dominantnem tipu, se razvijejo šele v določeni starosti. Torej, Huntingtonova koreja se klinično manifestira pri starosti 35-40 let, policistična ledvična bolezen se kaže tudi pozno. Zato se pri napovedovanju takšnih bolezni ne upoštevajo bratje in sestre, ki niso dosegli kritične starosti.
Prvi opis rodovnika z avtosomno prevladujočim tipom dedovanja anomalije pri ljudeh je bil podan leta 1905. Sledi prenos v več generacijah. brahidaktilija(s kratkimi prsti). Na sl. 6.26 prikazuje rodovnik s to anomalijo. Na sl. 6.27 prikazuje rodovnik z retinoblastomom v primeru nepopolne penetracije.
Rodovniki pri avtosomno recesivnem dedovanju. Recesivne lastnosti se fenotipsko pojavijo le pri homozigotih za recesivne alele. Te lastnosti praviloma najdemo pri potomcih fenotipsko normalnih staršev - nosilcev recesivnih alelov. Verjetnost pojava recesivnega potomstva je v tem primeru 25%. Če ima eden od staršev recesivno lastnost, bo verjetnost njegove manifestacije pri potomcih odvisna od genotipa drugega starša. Pri recesivnih starših bodo vsi potomci podedovali ustrezno recesivno lastnost (slika 6.28).

riž. 6.26. Rodovnik ( AMPAK) z avtosomno dominantnim tipom dedovanja (brahidaktilija - B)
Za rodovnike z avtosomno recesivnim tipom dedovanja je značilno, da se lastnost ne pojavlja v vsaki generaciji. Najpogosteje se recesivni potomci pojavljajo pri starših z dominantno lastnostjo, verjetnost takšnega potomstva pa se pojavi v tesno sorodnih zakonih, kjer sta oba starša lahko nosilca istega recesivnega alela, pridobljenega od skupnega prednika. Primer avtosomno recesivnega dedovanja je rodovnik družine s psevdohipertrofična progresivna miopatija, v katerih so pogoste tesno povezane poroke (slika 6.29). Opozarja na širjenje bolezni v zadnji generaciji horizontalno.

riž. 6.27. Rodovnik z retinoblastomom v primeru nepopolne penetracije

riž. 6.28. Verjetnost, da bi imeli potomce z recesivno lastnostjo
iz različnih zakonskih parov ( I-IV)
Rodovniki s prevladujočim X-vezanim dedovanjem lastnosti. Geni, ki se nahajajo na kromosomu X in nimajo alelov na kromosomu Y, so prisotni v genotipih moških in žensk v različnih odmerkih. Ženska prejme svoja dva kromosoma X in ustrezne gene od očeta in matere, moški pa svoj edini kromosom X podeduje le od matere. Razvoj ustrezne lastnosti pri moških določa en sam alel, ki je prisoten v njegovem genotipu, pri ženskah pa je posledica interakcije dveh alelnih genov. V zvezi s tem se v populaciji z različno verjetnostjo najdejo lastnosti, podedovane po X-vezanem tipu pri moških in ženskah.
S prevladujočim dedovanjem, vezanim na X, je ta lastnost pogostejša pri ženskah zaradi njihove večje sposobnosti sprejemanja ustreznega alela od očeta ali matere. Moški lahko to lastnost podedujejo le po materi. Ženske z dominantno lastnostjo jo prenašajo enako na svoje hčere in sinove, moški pa le na svoje hčere. Sinovi nikoli ne podedujejo prevladujoče lastnosti, povezane z X-om, od svojih očetov.

riž. 6.29. Rodovnik z avtosomno recesivno dedovanjem (psevdohipertrofična progresivna miopatija)
Primer te vrste dedovanja je rodovnik, opisan leta 1925 s folikularno keratozo - kožna bolezen, ki jo spremlja izguba trepalnic, obrvi, las na glavi (slika 6.30). Značilen je težji potek bolezni pri hemizigotnih moških kot pri ženskah, ki so najpogosteje heterozigoti.
Pri nekaterih boleznih opazimo smrt moških hemizigotov v zgodnjih fazah ontogeneze. Potem naj bodo v rodovniku med obolelimi le ženske, pri katerih je razmerje med prizadetimi hčerami, zdravimi hčerkami in zdravimi sinovi 1:1:1. Moške dominantne hemizigote, ki ne umrejo v zelo zgodnjih fazah razvoja, najdemo pri spontanih splavih ali med mrtvorojenimi otroki. Za takšne značilnosti dedovanja pri ljudeh je značilna pigmentna dermatoza.
Rodovniki z recesivnim X-vezanim dedovanjem lastnosti. Značilna lastnost rodovnikov s to vrsto dedovanja je prevladujoča manifestacija lastnosti pri hemizigotnih moških, ki jo podedujejo od mater z dominantnim fenotipom, ki so nosilke recesivnega alela. Praviloma to lastnost podedujejo moški skozi generacijo od dedka po materi do vnuka. Pri ženskah se kaže le v homozigotnem stanju, za katerega se verjetnost poveča s tesno povezanimi porokami.
Najbolj znan primer recesivnega X-vezanega dedovanja je hemofilija. Dedovanje hemofilije tipa A je predstavljeno v rodovniku potomcev angleške kraljice Viktorije (slika 6.31).

riž. 6.30. Rodovnik z dominantnim tipom dedovanja, vezanim na X (folikularna keratoza)

riž. 6.31. Rodovnik z X-vezanim recesivnim dedovanjem (hemofilija tipa A)
Drug primer dedovanja te vrste je barvna slepota - določena oblika motnje barvnega vida.
Rodovniki pri dedovanju, vezanem na Y. Prisotnost Y-kromosoma samo pri moških pojasnjuje značilnosti Y-povezanega ali hollandskega dedovanja lastnosti, ki jo najdemo samo pri moških in se prenaša po moški liniji iz roda v rod od očeta do sina.

riž. 6.32. Rodovnik z Y-vezanim (holandskim) tipom dedovanja
Ena lastnost, katere dedovanje, povezano z Y pri ljudeh, je še vedno predmet razprave hipertrihoza ušesa, ali prisotnost dlak na zunanjem robu ušesa. Domneva se, da poleg tega gena kratka roka Y kromosoma vsebuje gene, ki določajo moški spol. Leta 1955 so pri miši opisali transplantacijski antigen, vezan na Y, imenovan HY. Morda je to eden od dejavnikov spolne diferenciacije moških spolnih žlez, katerih celice imajo receptorje, ki vežejo ta antigen. Antigen, vezan na receptor, aktivira razvoj spolnih žlez moškega tipa (glejte poglavja 3.6.5.2; 6.1.2). Ta antigen je med evolucijo ostal skoraj nespremenjen in se nahaja v telesu številnih živalskih vrst, vključno z ljudmi. Tako je dedovanje sposobnosti razvoja spolnih žlez po moškem tipu določeno s holandskim genom, ki se nahaja na Y kromosomu (slika 6.32).
dvojna metoda
Ta metoda je sestavljena iz preučevanja vzorcev dedovanja lastnosti v parih enojajčnih in dizigotnih dvojčkov. Leta 1875 ga je predlagal Galton, da bi ocenil vlogo dednosti in okolja pri razvoju človekovih duševnih lastnosti. Trenutno se ta metoda široko uporablja pri preučevanju dednosti in variabilnosti pri ljudeh, da se ugotovi relativna vloga dednosti in okolja pri nastanku različnih znakov, tako normalnih kot patoloških. Omogoča vam, da ugotovite dedno naravo lastnosti, določite penetracijo alela, ocenite učinkovitost delovanja na telo nekaterih zunanji dejavniki (zdravila, usposabljanje, izobraževanje).
Bistvo metode je primerjati manifestacijo lastnosti v različne skupine dvojčkov na podlagi podobnosti ali razlik v njihovih genotipih. monozigotni dvojčki, ki se razvijejo iz enega samega oplojenega jajčeca, so genetsko identični, saj imajo 100 % skupnih genov. Zato je med monozigotnimi dvojčki visok odstotek skladni pari, pri katerem se lastnost razvije pri obeh dvojčkih. Primerjava monozigotnih dvojčkov, vzgojenih v različnih pogojih postembrionalnega obdobja, omogoča ugotavljanje znakov, pri katerih imajo okoljski dejavniki pomembno vlogo. Po teh znakih je med dvojčki neskladje, tiste. razlike. Nasprotno, ohranjanje podobnosti med dvojčki kljub razlikam v pogojih njihovega obstoja kaže na dedno pogojenost lastnosti.
Primerjava parne skladnosti po dana lastnost pri genetsko identičnih enojajčnih in dvoličnih dvojčkih, ki imajo v povprečju približno 50 % skupnih genov, omogoča objektivnejšo presojo vloge genotipa pri oblikovanju lastnosti. Visoka skladnost pri parih enojajčnih dvojčkov in bistveno manjša skladnost pri parih dvozigotnih dvojčkov kažeta na pomen dednih razlik v teh parih za določanje lastnosti. Podobnost indeksa skladnosti pri mono- in dizigotnih dvojčkih kaže na nepomembno vlogo genetskih razlik in na odločilno vlogo okolja pri nastanku znaka ali razvoju bolezni. Bistveno različne, a precej nizke stopnje skladnosti v obeh skupinah dvojčkov omogočajo presojo dedne nagnjenosti k nastanku lastnosti, ki se razvije pod vplivom okoljskih dejavnikov.
Ugotavljanje relativne vloge dednosti in okolja pri razvoju različnih patoloških stanj omogoča zdravniku, da pravilno oceni stanje in izvaja preventivne ukrepe v primeru dedne nagnjenosti k bolezni ali izvaja pomožno terapijo v primeru dedne pogojenosti.
Težave metode dvojčkov so povezane, prvič, z relativno nizko rodnostjo dvojčkov v populaciji (1:86-1:88), kar otežuje izbiro zadostnega števila parov s to lastnostjo; drugič, z identifikacijo monozigotnosti dvojčkov, kar je zelo pomembno za pridobitev zanesljivih zaključkov.
Za identifikacijo monozigotnosti dvojčkov se uporabljajo številne metode. 1. Polisimptomatska metoda za primerjavo dvojčkov po številnih morfoloških značilnostih (pigmentacija oči, las, kože, oblika las in značilnosti lasne linije na glavi in telesu, oblika ušes, nosu, ustnic, nohtov, telesa, vzorci prstov). 2. Metode, ki temeljijo na imunološki identiteti dvojčkov za eritrocitne antigene (ABO, MN, Rhesus sistemi), za serumske beljakovine (γ-globulin). 3. Najbolj zanesljivo merilo za monozigotnost je transplantacijski test z navzkrižnim cepljenjem kože dvojčkov.
Kljub zahtevnosti metode dvojčkov in možnosti napak pri določanju monozigotnosti dvojčkov, je zaradi visoke objektivnosti ugotovitev ena izmed široko uporabljenih metod genetskih raziskav pri ljudeh.
Sodobna klinična medicina ne more več brez genetskih metod. Za preučevanje dednih lastnosti pri ljudeh se uporabljajo različne biokemične, morfološke, imunološke in elektrofiziološke metode. Zahvaljujoč napredku genetskih tehnologij je mogoče laboratorijske genetske diagnostične metode izvajati na majhni količini materiala, ki ga lahko pošljemo po pošti (nekaj kapljic krvi na filtrirnem papirju ali celo na eni celici, vzeti v zgodnji fazi razvoja ( NP Bočkov, 1999) (slika 1.118).
riž. 1.118. M. P. Bochkov (rojen 1931)
Pri reševanju genetskih problemov se uporabljajo naslednje metode: genealoška, dvojčka, citogenetska, hibridizacija somatskih celic, molekularno genetske, biokemijske, dermatoglifne in palmoskopske metode, populacijska statistika, sekvenciranje genoma itd.
Genealoška metoda za preučevanje človekove dednosti
Glavna metoda genetske analize pri ljudeh je sestavljanje in preučevanje rodovnika.
Genealogija je rodoslovje. Genealoška metoda je metoda rodovnikov, ko se v družini izsledi lastnost (bolezen), ki kaže na družinske vezi med člani rodovnika. Temelji na temeljitem pregledu družinskih članov, sestavljanju in analizi rodovnikov.
To je najbolj univerzalna metoda za preučevanje človeške dednosti. Vedno se uporablja, če obstaja sum na dedno patologijo, pri večini bolnikov vam omogoča, da ugotovite:
dedna narava lastnosti;
Vrsta dedovanja in penetracija alelov;
Narava povezave genov in izvajanje preslikave kromosomov;
Intenzivnost procesa mutacije;
Dešifriranje mehanizmov interakcije genov.
Ta metoda uporablja v genetskem svetovanju.
Bistvo genealoške metode je vzpostavitev družinskih vezi, simptomov ali bolezni med bližnjimi in daljnimi, neposrednimi in posrednimi sorodniki.
Sestavljen je iz dveh stopenj: izdelava rodovnika in genealoška analiza. Preučevanje dedovanja lastnosti ali bolezni v določeni družini se začne pri subjektu, ki ima to lastnost ali bolezen.
Posameznik, ki ga genetik prvi opazi, se imenuje proband. Pretežno je bolnik ali nosilec raziskovalnih znakov. Otroci enega starševskega para se imenujejo probandovi bratje (bratje - sestre). Potem gredo k njegovim staršem, potem k bratom in sestram staršev in njihovih otrok, potem k starim staršem itd. Pri sestavljanju rodovnika naredite kratke opombe o vsi od družinskih članov, njegove družinske vezi s probandom. Rodovniško shemo (slika 1.119) spremljajo simboli pod sliko in se imenuje legenda.

riž. 1.119. Rodovnik družine, kjer se podeduje siva mrena:
bolniki s to boleznijo so družinski člani I - 1, I In - 4, III - 4,
Z uporabo genealoške metode je bilo mogoče ugotoviti naravo dedovanja hemofilije, brahidaktilije, ahondroplazije itd. Široko se uporablja za razjasnitev genetske narave patološkega stanja in napovedovanje zdravja potomcev.
Metode sestavljanja rodovnikov, analiza. Sestavljanje rodovnika se začne s probandom - oseboki se je obrnil na genetika ali zdravnika in vsebuje lastnost, ki jo je treba preučiti pri sorodnikih po očetovski in materini liniji.
Pri sestavljanju rodoslovnih tabel uporabljajo konvencije, ki jih je leta 1931 predlagal G. Yust (slika 1.120). Rodovniške figure so postavljene vodoravno (ali vzdolž krog), ena vrstica vsako generacijo. Na levi strani je vsaka generacija označena z rimskimi številkami, posamezniki v generaciji pa so označeni v arabščini od leve proti desni in od zgoraj navzdol. Poleg tega je najstarejša generacija postavljena na vrh rodovnika in je označena s številko i, najmanjša pa na dnu rodovnika.

riž. 1.120. Simboli, ki se uporabljajo pri sestavljanju rodovnikov.
Bratje in sestre v zvezi z rojstvom najstarejšega se nahajajo na levi strani. Vsak član rodovnika ima svojo kodo, npr. II - 4, II in - 7. Zakonski par v rodovniku je označen z isto številko, vendar z malo črko. Če eden od zakoncev ni poročen, podatki približno sploh ni dano. Vsi posamezniki so postavljeni strogo po generacijah. Če je rodovnik odličen, potem različne generacije niso razporejene v vodoravne vrste, ampak v koncentrične.
Po sestavi rodovnika se mu priloži pisna razlaga - legenda rodovnika. Naslednji podatki se odražajo v legendi:
Rezultati kliničnega in postkliničnega pregleda probanda;
Informacije o osebni preiskavi sorodnikov proband;
Primerjava rezultatov osebnega pregleda probanda po anketi njegovih sorodnikov;
Pisni podatki o sorodnikih, ki živijo na drugem območju;
Zaključek glede vrste dedovanja ali lastnosti bolezni.
Pri sestavljanju rodovnika se ne smete omejiti le na anketo sorodnikov - to ni dovolj. Nekateri od njih predpišejo popoln klinični, postklinični ali specialni genetski pregled.
Namen genealoške analize je ugotoviti genetske vzorce. Za razliko od drugih metod je treba genealoško raziskavo zaključiti z genetsko analizo njenih rezultatov. Analiza rodovnika omogoča sklepanje o naravi lastnosti (dedna ali ne), nazivu, dedovanju (avtosomno dominantno, avtosomno recesivno ali spolno vezano), zigotnosti probanda (homo - ali heterozigota), stopnji penetracije in ekspresivnosti preučevanega gena
Značilnosti rodovnikov z različnimi vrstami dedovanja: avtosomno dominantno, avtosomno recesivno in členjeno. Analiza rodovnikov kaže, da vse bolezni, ki jih določa mutantni gen, ustrezajo klasičnim zakoni Mendel za različne vrste dedovanja.
Po avtosomno dominantnem načinu dedovanja prevladujoči geni fenotipsko manifestirajo v heterozigotnem stanju, zato njihova določitev in narava dedovanja ne povzročata težav.
1) pri vsaki prizadeti osebi je eden od staršev bolan;
2) pri prizadeti osebi, ki je poročena z zdravo žensko, je v povprečju polovica otrok bolnih, druga polovica pa zdrava;
3) zdravi otroci prizadetega starša imajo zdrave otroke in vnuke;
4) moški in ženske so prizadeti enako pogosto;
5) bolezen se mora manifestirati v vsaki generaciji;
6) prizadeti heterozigotni posamezniki.
Primer avtosomno prevladujoče vrste dedovanja je lahko narava dedovanja šesteroprstega (velikoprstnega). Šestprsti udi so precej redek pojav, vendar se vztrajno ohranjajo v mnogih generacijah nekaterih družin (slika 1.121). Bagatopalia se pri potomcih dosledno ponavlja, če je vsaj eden od staršev bugatopalia, in je odsoten v primerih, ko so okončine normalne pri obeh starših. Pri potomcih bogatih staršev je ta lastnost v enakem številu prisotna pri fantih in deklicah. Delovanje tega gena v ontogenezi se pojavi precej zgodaj in ima visoko penetracijo.

riž. 1.121. Rod z avtosomno prevladujočim tipom dedovanja.
Pri avtosomno dominantnem dedovanju je tveganje za razvoj bolezni pri potomcih, ne glede na spol, 50%, vendar so manifestacije bolezni v določeni meri odvisne od penetracije.
Analiza porekel kaže, da syndactyly, Marfan bolezen, ahondroplazij, brachydactyly, Osler je hemoragična teleangiektazija, hemachromatosis, hiperbilirubinemija, hiperlipoproteinemija, različne dysostoses, marmor bolezen, nepopolni osteogeneze, Recklinghausen je nevrofibromatoza, otoskleroza, Peltzius-Merzbacherjeva bolezen, pelgirivirus levkociti, periodično adynamia, perniciozna anemija, polidaktilija, akutna intermitentna porfirija, dedna ptoza, idiopatska trombocitopenična purpura, talasemija, tuberozna skleroza, favizem, Charcot-Mariejeva bolezen, Sturge-Weberjeva bolezen, večkratne eksostoze, ektotopija leče, LO17 in drugi.
Glede na avtosomno recesivno dedovanje se recesivni geni fenotipsko pojavljajo le v homozigotnem stanju, kar otežuje tako identifikacijo kot preučevanje narave dedovanja.
Za to vrsto dedovanja so značilni naslednji vzorci:
1) če se je bolan otrok rodil fenotipsko normalnim staršem, potem so starši nujno heterozigoti;
2) če so bili prizadeti bratje in sestre rojeni iz tesno povezane zakonske zveze, je to dokaz recesivnega dedovanja bolezni;
3) če se poročijo z bolno recesivno boleznijo in genotipsko normalno osebo, bodo vsi njihovi otroci heterozigoti in fenotipsko zdravi;
4) če je zakon bolan in heterozigot, potem bo prizadeta polovica njihovih otrok in pol - heterozigoten;
5) če se dva bolnika poročita zaradi iste recesivne bolezni, bodo vsi njihovi otroci bolni.
6) moški in ženske zbolijo enako pogosto:
7) heterozigoti so fenotipsko normalni, vendar so nosilci ene kopije mutantnega gena;
8) prizadeti posamezniki so homozigotni, njihovi starši pa so heterozigotni nosilci.
Analiza rodovnikov kaže, da se fenotip detekcije recesivnih genov pojavlja le v tistih družinah, kjer imajo ti geni oba starša vsaj v heterozigotnem stanju (slika 1.122). Recesivni geni v človeški populaciji ostajajo neodkriti.

riž. 1.122. Rod z avtosomno recesivno vrsto dedovanja.
Vendar pa se v zakonih med ožjimi sorodniki ali v izoliranih (majhnih skupinah ljudi), kjer zakonske zveze potekajo po tesnih družinskih vezah, poveča izražanje recesivnih genov. V takih pogojih se verjetnost prehoda v homozigotno stanje in fenotipska manifestacija redkih recesivnih genov močno poveča.
Ker ima večina recesivnih genov negativen biološki pomen in povzroča zmanjšanje vitalnosti ter pojav različnih moškosti in dednih bolezni, imajo sorodne poroke izrazito negativen značaj za zdravje potomcev.
Dedne bolezni se pretežno prenašajo avtosomno recesivno, otroci heterozigotnih staršev lahko podedujejo bolezni v 25 % primerov (s popolno penetracijo). Glede na to, da je popolna penetracija redka, je manjši tudi odstotek dedovanja bolezni.
Avtosomna recesivna dedna: agammaglobulіpemіya, agranulocitoza, alkaptonurіya, albinizem (. Slika 1,123), Amavrotichna idiot amіnoatsidurії, anemija, avtoimunska hemolitična anemija hipokromna mіkrotsitarna, anencefalija, galaktozemije, hermafroditizem (slika 1,124) gepagotserebralna distrofija, Gaucherjeva bolezen, єvnuhoїdizm, myxedema, srpastih celična anemija, fruktozurija, barvna slepota(L. O. Badalyan et al., 1971).

riž. 1.123. - Dedovanje po avtosomno recesivnem tipu. albinizem.

riž. 1.124. Avtosomno recesivno dedovanje. Hermafroditizem.
Številne bolezni se dedujejo po X-kromosomskem (spolno vezanem) tipu, ko je mati nosilka mutantnega gena, polovica njenih sinov pa je bolnih. Obstaja X-povezano dominantno X-povezano recesivno dedovanje.
Rod X-povezanega dominantnega dedovanja (slika 1.125). Za to vrsto dedovanja je značilno:
1) Prizadeti moški prenašajo svojo bolezen na svoje hčere, ne pa na svoje sinove;
2) prizadete heterozigotne ženske prenašajo bolezni na polovico svojih otrok, ne glede na njihov spol;
3) Prizadete homozigotne samice prenašajo bolezen na vse svoje otroke.
Ta vrsta dedovanja ni pogosta. Bolezen pri ženskah ni tako huda kot pri moških. Precej težko je razlikovati sebe X-povezano dominantno in avtosomno dominantno dedovanje. Uporaba novih tehnologij (DNK sond) pomaga natančneje določiti vrsto dedovanja.

riž. 1.125. X-vezano dominantno dedovanje.
Rodovid X-vezano recesivno dedovanje (slika 1.126). Za to vrsto so značilni takšni vzorci dedovanja:
1) skoraj vsi prizadeti so moški;
2) lastnost se prenaša preko heterozigotne matere, ki je fenotipsko zdrava;
3) prizadeti oče nikoli ne prenese bolezni na svoje sinove;
4) vse hčere bolnega očeta bodo heterozigotne nosilke;
5) ženska prenašalka bolezen prenese na polovico svojih sinov, nobena od hčera ne bo bolna, ampak pol hčere - nosilke dednega gena.

riž. 1.126. X-vezano recesivno dedovanje.
Več kot 300 lastnosti povzročajo mutantni geni, ki se nahajajo na kromosomu X.
Primer recesivnega dedovanja spolno vezanega gena je hemofilija. Bolezen je relativno pogosta pri moških in zelo redka pri ženskah. Fenotipsko zdrave ženske so včasih "nosilke" in ko so poročene z zdrav človek roditi sinove s hemofilijo. Takšne ženske so heterozigotne za gen, ki povzroča izgubo sposobnosti strjevanja krvi. Iz porok moških s hemofilijo z zdravimi ženskami se vedno rodijo zdravi sinovi in hčerke nosilke, iz porok zdravih moških z nosilkami pa je polovica sinov bolnih in polovica hčera nosilk. Kot že omenjeno, je to posledica dejstva, da oče svoj X kromosom prenese na svoje hčere, sinovi pa prejmejo samo Y -kromosom, ki nikoli ne vsebuje gena za hemofilijo, medtem ko njihov edini kromosom X prihaja od matere.
V nadaljevanju so navedene glavne bolezni, ki se dedujejo po recesivnem, spolno povezanem tipu.
Agamaglobulinemija, albinizem (nekatere oblike), hipokromna anemija, Wiskott-Aldrichov sindrom, Hutnerjev sindrom, hemofilija A, hemofilija B, hiperparatiroidizem, glikogenoza tipa VI, pomanjkanje glukoza-6-fosfat dehidrogenaze, sladkorna bolezen nefrotizioze bolezen Merzbacherja, periodična paraliza, pigmentoza retinitisa, psevdohipertrofična oblika miopatije, Fabryjeva bolezen, fosfatna sladkorna bolezen, Scholzova bolezen, barvna slepota (slika 1.127).

riž. 1.127. Test za določanje zaznavanja barv z Rabkinovimi tabelami.
genealoška metoda metoda preučevanja narave dedovanja določene lastnosti ali ocenjevanja verjetnosti njenega pojava v prihodnosti med člani preučevane družine, ki temelji na ugotavljanju družinskih vezi (rodovnik) in sledenju lastnosti med vsemi sorodniki.
1. Majhna medicinska enciklopedija. - M.: Medicinska enciklopedija. 1991-96 2. Prva pomoč. - M.: Velika ruska enciklopedija. 1994 3. Enciklopedični slovar medicinskih izrazov. - M.: Sovjetska enciklopedija. - 1982-1984.
Poglejte, kaj je "Genealoška metoda" v drugih slovarjih:
Metoda preučevanja narave dedovanja določene lastnosti ali ocenjevanja verjetnosti njenega pojava v prihodnosti med člani preučevane družine, ki temelji na ugotavljanju družinskih vezi (rodovnik) in sledenju lastnosti med vsemi sorodniki ... Veliki medicinski slovar
GENEALOŠKA METODA- [cm. genealogija] metoda preučevanja narave dedovanja določene lastnosti ali ocenjevanja verjetnosti njenega pojava v prihodnosti med člani preučevane družine; G. m. se uporablja pri preučevanju narave dedovanja lastnosti (na primer bolezni) ... Psihomotorika: Referenčni slovar
genealoška metoda- v človeški genetiki metoda analize rodovnikov. Uporablja se za preučevanje narave porazdelitve dednih lastnosti v družinah. V medicini se pogosteje uporablja za genetsko analizo različnih patoloških nepravilnosti ... Slovar psihogenetike
genealoška metoda- se nanaša na psihogenetske metode. Izvaja se študija podobnosti med sorodniki v različnih generacijah. To zahteva natančno poznavanje številnih značilnosti neposrednih sorodnikov po materini in očetovi liniji ter pokritost v največji možni meri ... ...
psihogenetska metoda- (metoda psi "genetike) metode, ki omogočajo ugotavljanje vpliva dednih dejavnikov in okolja na oblikovanje določenih duševnih značilnosti osebe (glej psihogenetika). Ti vključujejo: 1) metoda dvojčka je najbolj informativna; 2) metoda ... ... Velika psihološka enciklopedija
psihogenetika- (iz grškega genetikos, ki se nanaša na rojstvo, izvor) področje psihologije, ki meji na genetiko. Predmet P. je izvor individualnih psiholoških značilnosti človeka, razjasnitev vloge genotipa in okolja pri njihovem nastanku. P … Velika psihološka enciklopedija
Psihogenetika (grško psihična duša in grški izvor geneze) je veda o dednosti in variabilnosti duševnih in psihofizioloških lastnosti, ki je nastala na stičišču psihologije in genetike. V zahodni književnosti ... Wikipedia
Veja psihologije, ki uporablja genetske podatke in genealoško metodo. Predmet psihogenetike je interakcija dednosti in okolja pri oblikovanju interindividualne variabilnosti človeških psiholoških lastnosti (kognitivnih in ... ... Psihološki slovar
Odsek človeške genetike, namenjen preučevanju vloge dednih dejavnikov v človeški patologiji na vseh glavnih ravneh organizacije življenja od populacije do molekularne genetike. Glavni odsek M.g. predstavlja klinično genetiko, ... ... Medicinska enciklopedija
Veja genetike, ki je tesno povezana z antropologijo in medicino. G. ure pogojno delimo na antropogenetiko, ki proučuje dednost in spremenljivost normalnih znakov človeškega telesa, in medicinsko genetiko (glej Genetika ... ... Velika sovjetska enciklopedija
I Dednost je lastnost vseh organizmov, da v številnih generacijah zagotavlja kontinuiteto znakov in značilnosti razvoja, torej morfološko, fiziološko in biokemično organizacijo živih bitij in naravo njihovega posameznika ... .. . Medicinska enciklopedija
 Ali otrok potrebuje povodec za hojo?
Ali otrok potrebuje povodec za hojo?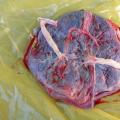 Kakšna je posteljica v telesu nosečnice?
Kakšna je posteljica v telesu nosečnice?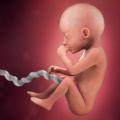 Sedmi mesec nosečnosti: razvoj otroka
Sedmi mesec nosečnosti: razvoj otroka